
Além da seleção natural ou a importância da evolução neutra.
A palavra “evolução” descreve o padrão arborescente de ancestralidade comum, a poderosa idéia de Darwin, que gera uma “hierarquia aninhada” com as mesmas características das árvores genealógicas de famílias humanas, estendendo a noção de parentesco à todas as espécies.
Entretanto, “evolução”, em seu sentido minimo (estando este significado implícito no sentido mais popular, comentado no parágrafo anterior), é simplesmente a mudança transgeracional de características hereditárias de uma população, algo que é maravilhosamente capturado pela expressão criada por Darwin, “descendência com modificação, antes que o termo “evolução” fosse empregado para descrever este processo. Em realidade, esta
mudança transgeracional depende de outros dois processos. O primeiro é a geração de variabilidade hereditária e o segundo é a sobrevivência e reprodução diferencial destas variantes. Por este motivo, a evolução é muitas vezes definida operacionalmente, por geneticistas de populações, como mudanças das freqüências alélicas (isto é, de variantes gênicas) ao longo das gerações em uma população.
Darwin, juntamente com o naturalista, Alfred Russell Wallace, contribuiu com outra idéia fundamental ao estudo da evolução biológica, a seleção natural. Este conceito surgiu em analogia a seleção artificial de características morfológicas e fisiológicas, levada a cabo por criadores de animais e cultivadores de plantas. O termo “seleção natural” descreve o processo de sobrevivência e reprodução diferencial dos indivíduos de uma população, quando este é afetado causalmente, diga-se de passagem, pela posse de determinadas características hereditárias, desta forma aumentando a proporção destas características nas gerações futuras da população em questão. Então, o destino da variabilidade hereditária (i.e., genética, criada por mutações) quando estas
alteram características, que envolvem as capacidades de sobrevivência e reprodução dos organismos que as possuem, é, parcialmente, determinado pela seleção natural.
Desta forma, caso os novos alelos, introduzidos em uma população por mutações, tornem seus portadores mais propensos a deixarem mais descendentes, aumentando sua aptidão, conferindo vantagens aos portadores destas características, estamos observando “seleção natural positiva”. Por outro lado, caso as mutações diminuam a aptidão de seus portadores, diminuindo suas chances de sobrevivência e reprodução (portanto, reduzindo sua representação nas próximas gerações), temos o que se chama de “seleção natural negativa
(ou purificadora)”. Em uma terceira situação, na qual o alelo só traga vantagem em heterozigose, observamos a “seleção balanceadora” que manterá ambas versões alélicas na população, mesmo que individualmente, em homozigose, estes mesmos alelos sejam deletérios e causem prejuízo aos indivíduos que os possuem.
No caso em que a seleção positiva está atuando, ao longo do tempo, ela elevará a freqüência do alelo que confere maior aptidão em uma dada população. Por fim, levando a substituição completa do alelo anterior pelo alelo mutante, o que se chama de fixação, ou seja, a freqüência do alelo chega a 100% naquela dada população.
Entretanto, ao analisarmos estes processos com mais atenção, notamos que para que ocorra seleção natural é preciso que os alelos envolvidos causem, pelo menos em parte, diferenciais reprodutivos, alterando a aptidão de seus portadores. Caso contrário não haverá seleção. Se isso não acontece, ainda assim, a evolução continua.
Se as populações, simplesmente, não continuarem crescendo indefinidamente, ao longo das gerações, as freqüências genéticas dos alelos tenderão a flutuar. Imagine, por exemplo, que os diversos alelos presentes em uma população confiram exatamente a mesma aptidão. Mesmo assim, apenas uma fração dos gametas darão origem a indivíduos adultos capazes de gerar seus próprios gametas e, assim, se reproduzir. Uma grande quantidade de alelos será perdida apenas pelo efeito de erro de amostragem no processo de reprodução. Este efeito será, quanto mais drástico, quanto menor o tamanho da população.
Este efeito é basicamente produzido por "erros de amostragem estatística" a cada geração, já que, da enorme gama de gametas produzidos, apenas uma fração diminuta contribuirá à proxima geração. A figiua abaixo ilustra o efeito desta dinamica de amostragem aleatória (clique nela para ver a animação).

Neste caso, quando as populações são pequenas, mesmo alelos que confiram vantagens adaptativas, aos seus portadores, podem ser perdidos nesta “loteria” reprodutiva. De forma complementar, alelos que confiram pequenas desvantagens, diminuindo a aptidão, podem ser mantidos e levados a fixação em uma população.
Este processo, chamado de deriva genética, proposto por Sewall Wrigth, é de extrema importância para evolução (Na figura ao lado, está ilustrado o efeito da deriva genética aleatória na probabilidade de fixação de alelos em relação ao tamanho da população.). Porém, apenas a partir dos anos sessenta, com os primeiros estudos sobre o relógio molecular e
a descoberta da grande diversidade bioquímica das proteínas nas populações biológicas, é que este fenômeno começou a ganhar sua devida atenção. Isto aconteceu, principalmente, através dos trabalhos do geneticista matemático japonês, Motoo Kimura, proponente da “teoria neutra da evolução molecular”. Pela primeira vez, os biólogos evolutivos possuíam uma elegante e robusta teoria “nula” para explicar a grande, e inesperada, diversidade molecular, além da constância do relógio molecular em muitas espécies diferentes. Esta teoria foi crucial na criação de testes para as hipóteses de seleção natural no nível molecular.
Os trabalhos de Kimura, e o artigo seminal de Jukes e King, iniciaram uma contenda científica conhecida como a “controvérsia selecionista vs. neutralista”. Este saudável e interessante debate, envolvendo os biólogos evolutivos, adentrou pelos anos 70 e 80, continuando de forma muito menos intensidade, até os dias de hoje. Este debate foi responsável por uma nova gama de abordagens matemáticas e teóricas que nos ajudam a compreender melhor a biologia evolutiva.
Na década de setenta, Kimura junto a uma colaboradora mais jovem, Tomoko Ohta, propuseram uma versão modificada da teoria neutra, a “teoria quase-neutra”. Nesta nova versão era dado muito mais atenção ao tamanho populacional e a possibilidade de que mutações ligeiramente deletérias pudessem ser levadas a fixação, por se comportar de forma “quase-neutra”, nestas situações.
As principais evidências que comprovam a importância da teoria neutra, foram sumarizado por Kimura, em um artigo de 1991:
Em seqüências de proteínas, substituições conservadoras de aminoácidos por aminoácidos com características físico-químicas semelhantes (ou seja, que causam poucas alteações bioquímicas sem
muito afetar a função protéica) ocorrem em maior freqüência que alterações mais radicais.Substituições sinônimas (i.e. aquelas que não alteram o aminoácido codificado) ocorrem quase sempre em uma taxa muito maior do que as substituições não sinônimas.
Seqüências não-codificadoras, como introns, evoluem em uma taxa mais alta, similar a de sítios sinônimos.
Pseudogenes, evoluem a uma taxa maior, semelhante a taxa da terceira posição de um códon.
Estas observações são extensamente corroboradas pelos dados genômicos e não se ajustam ao modelo selecionista tradicional.
Hoje a evolução neutra não pode mais ser ignorada, principalmente porque, ela esta na base dos testes para a detecção de seleção natural em genes. No entanto, hoje reconhece-se que a divisão das mutações em
deletérias, neuras e vantajosas é uma simplificação grosseira, havendo na realidade um continuum de efeitos fenotípicos que podemvariar de a cordo com o contexto, e, principalmente (como foi reconhecido pelos trabalhos de Kimura e Ohta), pelo tamanho efetivo da população.
Então, a importância da deriva genética vai ser proporcional a dois valores: (i) o tamanho efetivo da população, Ne,;e (ii) o coeficiente de seleção, s, que exprime a efeito fenotípico da mutação na aptidão do seu portador. Desta forma, mutações cujo valor de Nes for muito menor do que 1z, podem ser consideradas “efetivamente neutras”, tendo seu destino definido pelos efeitos estocásticos da deriva genética. Assim, quanto menor for Ne
maior será o efeito da deriva nos genes, mesmo que estes confiram vantagens adaptativas.
De acordo com a teoria neutra, a probabilidade de uma mutação neutra atingir a fixação é igual a sua freqüência na população, por isso, uma nova mutação tem probabilidade de 1/N (em uma população haplóide, mas o mesmo raciocínio é válido para populações diplóides), onde N é o tamanho da população. Então, se consideramos que a taxa de mutação em um determinado sítio é igual u, em uma população haplóide, a cada
geração, ocorrem Nu mutações. Assim, a taxa de substituições por geração (K) pode ser obtida simplesmente
ao multiplicar-se o número de mutações que ocorrem a cada geração ( Nu) pela sua probabilidade de fixação (1/N), o que nos dá, para sítios evoluindo de forma neutra:
K = Nu × 1/N = u
Porém, como a seleção natural altera a probabilidade de fixação de uma mutação (com a seleção positiva a aumentando, e a seleção negativa, a diminuindo, em relação as mutações neutras) espera-se que sítios afetados pela seleção positiva evoluam mais rápido (K>u), e, os afetados pela seleção negativa, evoluam mais
devagar (K<u) do que sítios neutros. Muitos dos testes para detectar a seleção natural baseiam-se nestas expectativas. Porém, é sempre preciso um pouco de cuidado, já que outros processos podem influenciar K, como a “conversão enviesada gênica” (BGC, Biased Gene conversion), assim como expansões populacionais e relaxamento da seleção negativa. Por isso, estes processos também devem ser levados em consideração. Portanto, em condições ideais, é importante que se tenha uma hipótese ecológico-funcional, ou bioquímica, para reforçar a hipótese seletiva, permitindo testar e estimar os ganhos em aptidão das variantes envolvidas.
Kimura e os defensores do modelo neutro de evolução jamais negaram a existência ou, mesmo, a importância da seleção natural, sobretudo para a evolução fenotípica. Apenas enfatizavam a maior importância relativa da fixação de alelos, seletivamente neutros, por deriva genética, na evolução molecular. Porém, nas últimas décadas um papel mais positivo para a deriva genética e outros processos estocásticos, tem sido ativamente investigado. Entre estes processos não adaptativos, podemos citar a deriva genômica em famílias multi-gênicas, como a dos receptores olfativos em primatas, e o aumento do tamanho e complexificação dos genomas, e da estrutura dos genes, nos eucariontes, por conta do acumulo de mutações efetivamente neutras propiciado pela diminuição no tamanho efetivo das populações através da árvore da vida.
A evolução dos genomas tem merecido especial atenção. Os genomas variam dramaticamente em relação ao tamanho (variando em 4 ordens de magnitude apenas nos eucariontes) e estrutura, com os genomas de procariontes sendo bastante compactos, densos em seqüências codificadoras, em marcante contraste aos genomas de eucariontes, repletos de elementos não codificadores, como seqüências inter-genicas, introns, regiões UTR e muito, mas muitos mesmo, transposons. Isto resulta em um padrão geral no qual, a grosso modo, a complexidade e tamanho dos genomas aumentam na passagem dos procariontes, para os eucariontes unicelulares e destes para os eucariontes multicelulares, como animais e plantas. Um crescente número de estudos que compilam os dados genômicos de espécies representantes dos três domínios, baseados em princípios de genética de populações muito bem estabelecidos, tem sugerido um modelo teórico capaz de explicar esta situação. O grupo de Michael Lynch, da Universidade de Indiana, vem propondo que muitas das
características genômicas são resultado de processos não adaptativos, muitos de natureza estocásticas baseados nas grandes diferenças nos tamanhos efetivos das populações (Ne) que tendem a decrescer drasticamente, na passagem de procariontes para eucariontes e de unicelulares para pluricelulares.
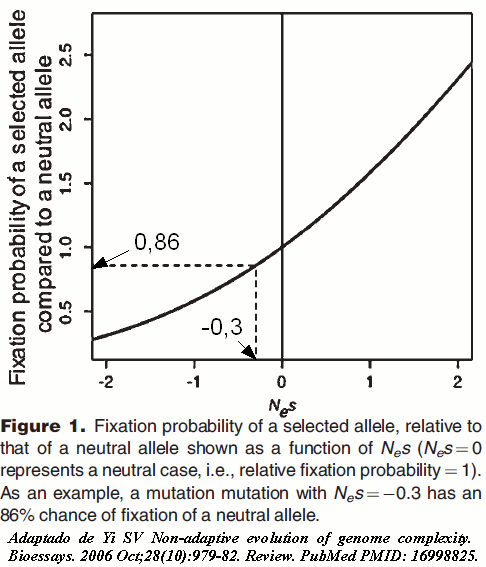
A taxa de deriva genética é proporcional a 1/Ne; e o efeito conjunto da seleção natural e da deriva genética em uma nova população, pode ser aproximado pela probabilidade de fixação de um novo alelo selecionado, relativamente a de um alelo neutro, o que dado por 4Nes/1 -e -4Nes. Assim o destino de uma nova mutação é determinado pelo seu efeito seletivo, relativizado pelo efeito do tamanho da população efetiva em que ele é introduzido. Portanto, um mutação que cause uma pequena desvantagem (por exemplo, s = -10 -5), caso surja em uma população com Ne=106, praticamente não terá chance de ser fixada. Não obstante, esta mesma mutação, caso a população tivesse Ne.=30000, ou seja Nes=-0,3, a chance de fixação desta mutação, ligeiramente desvantajosa, estaria por volta de 86% da chance de fixação de uma mutação neutra. Esta mutação se comportaria de forma muito semelhante a uma mutação neutra. Então, como já vimos,para mutações com |Nes| << 1, a deriva genética passa a ter uma importância substancial em seu destino. Por isso, Lynch propõe que mutações que satisfaçam |4Nes| < 1, cuja probabilidade de fixação seja 88% da de uma mutação neutra, sejam consideradas efetivamente neutras.
Na figura 1, em seguida, podemos observar como varia a probabilidade de fixação de um alelo selecionado em relação a um neutro, com o aumento do valor de Nes (no qual Nes=0, representa o caso da neutralidade, ou seja, probabilidade de fixação igual a 1). Estas considerações nos levam a perceber que o número de mutações
efetivamente neutras variará em função do tamanho populacional, como ilustrado na figura 2. Assim, a complexidade da arquitetura genômica de eucariontes, sobretudo dos multicelulares, é resultado da acumulação de mutações que cairiam na categoria de “efetivamente neutras”. Os genomas de espécies com pequenos Ne proveriam um ambiente genético adequado para o acúmulo de mutações quase neutras, como conseqüência da redução na eficiência da seleção purificadora, o que é corroborado pela diminuição nos tamanhos das populações efetivas através da “árvore da vida”. Assim, a complexidade genômica correlaciona-se negativamente com Ne.
O modelo de Lynch é também conformado pelo fato de que muitas das mutações, associadas ao ganho de complexidade estrutural nos genes e criação de redes regulatórias, seriam, pelo menos, ligeiramente deletérias, como ocorre com o acréscimo de introns (que precisa de cerca 20 a 40 pb, para que possam ser adequadamente reconhecidos e editados pelos spliceossomas) e regiões UTR (untranslated regions). A compilação de grande quantidade de dados de genomas dos três domínios revelam que a diversidade de sítios neutros (πs) que é equivalente à 4Neμ (onde μ é a taxa de mutação neutra) em populações diplóides em equilíbrio. Assim, como cada um dos sítios intrônicos podem sofrer mutações, que podem impedir seu correto funcionamento e processamento, é possível aproximar esta desvantagem seletiva, na inserção de um novo intron, por s=n μ, onde μ é a taxa de mutação por sítio. Então, para n=20, o valor crítico seria de 4Neμ= πs<0.05. Este valor é completamente compatível com as estimativas para sítios silenciosos (lembrando-se que |4Nes| < 1 pode ser considerados efetivamente neutros) em animais e plantas. Nestas espécies, que exibem grande quantidades de introns, este valor está bem a baixo deste ponto crítico. Porém, ainda mais interessante, é o fato de que o ajuste do dados da distribuição de introns entre procariontes e eucariontes unicelulares é excelente ao usarmos a hipótese de que πs= 0.05 represente o ponto crítico. Assim a redução da eficiência da seleção negativa, devido as pequenas populações, levaria a acumulação de introns, podendo levar a complexificação genômica. Além disso, algo muito semelhante seria também verdadeiro para o acúmulo de regiões UTR e para a modularização dos sistemas de regulação gênica. Tudo muito bem amparado pela teoria de genética de populações e a enorme gama de dados empíricos coletados pelo grupo de Lynch.

O modelo de Lynch explica os padrões gerais vistos na transição dos procariontes para os eucariontes e da unicelularidade para a multicelularidade, mas como o próprio Lynch admite, muitos dos detalhes de seu modelo se mostrarão errados. No entanto, a perspectiva geral parece ser muito robusta e oferece uma sólida e elegante hipótese a ser testadas. Resultados preliminares, ao analisar-se dados de herbívoros e carnívoros, parecem destoar das previsões do modelo de Lynch (ainda que o modelo tenha sido criado para explicar padrões bem mais gerais), mas estudos em peixes de
nadadeiras com arcos raiados, controlando para o metabolismo e tamanho corporal (dois outros fatores que podem confundir as análises e explicar o aumento do tamanho genômico, são compatíveis com as análises de Lynch.
A idéia de de que processos não adaptativos sejam os responsáveis por permitir a complexificação genômica, refinamentos na arquitetura dos genes e modularização das redes de controle genético, não exclui o papel da seleção natural, no ajuste e “aperfeiçoamento” destes sistemas, ou desvios desta tendências gerais em grupos particulares de eucariontes. Esta interação entre seleção e processos estocásticos, como mutação e deriva, especialmente em espécies
com substancial redução no tamanho das populações efetivas, é que parece ser a chave para a compreensão da evolução da complexidade e diversidade.
Referências:
Este artigo usou como fontes principais o ótimo artigo do portal scitable, de autoria de Laurent Duret, e uma esclarecedora revisão dos artigos de Michael Lynch, esrcrita por Soojin V. Yi, publicada no BioEssays,de onde foram retiradas as duas figuras usadas neste texto:
Duret, L. (2008) Neutral theory: The null hypothesis of molecular evolution. Nature Education 1(1)
Yi SV. Non-adaptive evolution of genome complexity. Bioessays. 2006 Oct;28(10):979-82. Review. PubMed PMID: 16998825.
Referências adicionais recomendadas:
Gregory TR, Witt JD. Population size and genome size in fishes: a closer look. Genome. 2008 Apr;51(4):309-13. PubMed PMID: 18356967.
King JL, Jukes TH. Non-Darwinian evolution. Science. 1969 May 16;164(881):788-98. PubMed PMID: 5767777.
Kimura, M. "Evolutionary Rate at the Molecular Level," Nature (1968), 217: 624-26.
Kimura M. Recent development of the neutral theory viewed from the Wrightian tradition of theoretical population genetics. Proc Natl Acad Sci U S A. 1991 Jul 15;88(14):5969-73. PubMed PMID: 2068072; PubMed Central PMCID: PMC52003.
Lynch M. The origins of eukaryotic gene structure. Mol Biol Evol. 2006 Feb;23(2):450-68. Epub 2005 Nov 9. PubMed PMID: 16280547.
Lynch M. The evolution of genetic networks by non-adaptive processes. Nat Rev Genet. 2007 Oct;8(10):803-13. PubMed PMID: 17878896.
Ohta, Tomoko; John H. Gillespie (1996). "Development of Neutral and Nearly Neutral Theories". Theoretical Population Biology 49 (2): 128–42. doi:10.1006/tpbi.1996.0007. , pp 130-131
Nei M, Rooney AP. Concerted and birth-and-death evolution of multigene families. Annu Rev Genet. 2005;39:121-52. Review. PubMed PMID: 16285855; PubMed Central PMCID: MC1464479.
Nei, M. (2005) Selectionism and neutralism in molecular evolution. Mol. Biol. Evol. 22:2318-2342.
Nei, M. (2006) Selectionism and neutralism in molecular evolution. Erratum Mol. Biol. Evol. 23:1095.
Stoltzfus A. On the possibility of constructive neutral evolution. J Mol Evol. 1999 Aug;49(2):169-81. PubMed PMID: 10441669.
Yi S, Streelman JT. Genome size is negatively correlated with effective population size in ray-finned fish. Trends Genet. 2005 Dec;21(12):643-6. Epub 2005 Oct 5. PubMed PMID: 16213058.
Livros recomendados:
Kimura, Motoo. 1983. The neutral theory of molecular evolution. Cambridge
Michael Lynch 2007 The Origins of Genome Architecture Sinauer
Transcrição da palestra de Lynch no Brasil:
http://www.revistapesquisa.fapesp.br/pdf/revolucao_genomica/lynch.pdf
Video da palestra de Lynch:
http://revistapesquisa.fapesp.br/?art=4977&bd=2&pg=1&lg=
Mais links de interesse:
http://authors.library.caltech.edu/5456/1/hrst.mit.edu/hrs/evolution/public/neutraltheory.html
http://authors.library.caltech.edu/5456/1/hrst.mit.edu/hrs/evolution/public/nearlyneutral.html
Crédito das imagens:
MARTIN KRZYWINSKI / SCIENCE PHOTO LIBRARY
NEMO RAMJET / SCIENCE PHOTO LIBRARY
Figuras sobre deriva genética:
Professor Marginalia através da commons.wikimedia.org.
{kind=link}