
Casais gays e formigas: a evolução pode explicar o altruísmo social?


Que semelhança notável há entre as formigas e a adoção de crianças por casais gays?
Não, não é que ambos causam urticária em muita gente. É que, ainda que haja controvérsia sobre quem pode adotar uma criança, não é surpresa que nós como seres sociais cuidemos dos filhos biológicos dos outros - assim como as formigas fazem.
A maioria das formigas que estão num formigueiro jamais terão descendentes. As operárias, todas filhas de uma mesma rainha, podem ter diferentes funções em diferentes castas, todas trabalhando de forma a manter ou expandir o formigueiro. Há inclusive as operárias que são babás, cuidam dos ovos e das jovens ninfas.

Um caso extremo está num tipo de formiga operária da espécie Myrmecocystus mexicanus, que regurgita o néctar guardado em seu abdômen expandido para alimentar suas irmãs em tempos de seca.
Já os casais humanos de orientação homossexual, incapazes de gerar descendentes biológicos por esta via, aparecem nas notícias todos os dias expressando o desejo de adotar crianças.
Pensando a seleção natural como a reprodução e sobrevivência diferenciais entre indivíduos, este mecanismo principal de mudança evolutiva proposto por Darwin parece ser insuficiente para explicar o comportamento social de "generosidade" das formigas e dos humanos. Se o que importa é ser mais apto (deixar mais descendentes), então parece que a evolução deveria gerar sempre comportamentos "egoístas", não comportamentos "altruístas" como esses.
Se todos fossem egoístas, a sociedade ruiria, tanto a das formigas quanto a dos humanos. A característica em comum entre humanos e formigas, que pode ser chamada de "eussocialidade" (do grego "eu", verdadeiro), representa então um problema a mais a ser resolvido pela teoria da evolução para explicar nossas origens.
A cooperação, então, é um elemento sine qua non do comportamento social. Como ela surgiu?
A teoria da seleção de parentesco era a mais badalada para explicar a origem deste comportamento até hoje. Esta teoria, desenvolvida por J. B. S. Haldane em 1955 e W. D. Hamilton em 1964, está fundada na noção de "aptidão inclusiva" e, como diz o nome, ela leva em conta o grau de parentesco. É um fato notável que a cooperação das formigas, por exemplo, é para com suas irmãs.
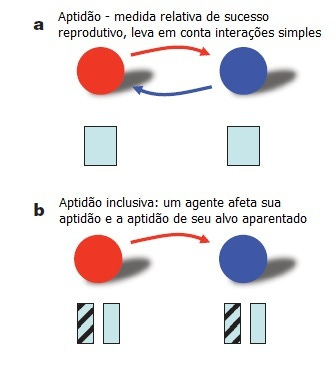
Atenção para o conceito: a aptidão inclusiva seria a soma entre o efeito da cooperação sobre a aptidão do agente (por exemplo a formiga doadora de néctar) e o efeito da cooperação sobre a aptidão do beneficiado (por exemplo as formigas nutridas pelo néctar) mutiplicada pelo parentesco entre o agente e o beneficiado.
Lembre: se estamos falando em mais apto e menos apto, estamos falando em seleção natural. O que este conceito veicula é que a seleção natural pode agir num tipo diferente de aptidão. E esta aptidão não é do indivíduo, é daquilo que o faz parente de outros indivíduos: os genes.
É por isso que o livro de divulgação "O Gene Egoísta", de Richard Dawkins, dedica-se a explicar o altruísmo e também a defender a noção de que é o gene a unidade sobre a qual a seleção natural age, e também seu nível principal de atuação.
A teoria de Haldane e Hamilton prevê que a evolução dará origem à cooperação entre organismos quanto mais aparentados eles forem. Hamilton, como bom cientista tradicional, tratou de expressar esta ideia matematicamente.
A cooperação será favorecida pela seleção natural sempre que o grau de parentesco R, que é a fração de genes compartilhados, for maior que a razão entre o custo c e o benefício b:
R > c / b
Logo, para irmãos, com R = 1/2, o altruísmo é um resultado esperado da evolução sempre que o custo para o altruísta for mais de duas vezes inferior ao benefício para seus irmãos. No caso de primos em primeiro grau, com R =1/8, o benefício teria de ser mais de oito vezes superior ao custo.
Esta teoria fez bastante sucesso porque explica muito bem a eussocialidade de organismos haplodiploides como as formigas e as abelhas; em que os sexos são determinados pelo número de cromossomos dos organismos, sendo os machos originados de ovos não fecundados com "n" cromossomos (machos haploides) e as fêmeas de ovos fecundados "2n" (diploides). Este ciclo implica que o R para as fêmeas tem o valor exorbitante de 3/4 - e todas as operárias de um formigueiro, que trabalham de forma abnegada para o "bem comum", são fêmeas!
Tudo bem fechadinho e plausível, não é? Mas, como dizia o filósofo Immanuel Kant, "a experiência sem teoria é cega, mas a teoria sem experiência é mera brincadeira intelectual".

Seria a teoria da seleção de parentesco mera brincadeira intelectual? Segundo um artigo de Martin Nowak, Corina Tarnita e o famoso sociboiólogo Edward O. Wilson, publicado este mês na revista Nature, parece que sim.
Os autores lembram que existem espécies eussociais, como os cupins e os ratos-toupeiras, que não são haplodiploides, e que há famílias inteiras de himenópteros (ordem das formigas e abelhas), com até 70 mil espécies, que são haplodiploides mas não eussociais. Então, ao menos a hipótese de associação entre haplodiploidia e eussocialidade, decorrente da teoria da seleção de parentesco (TSP), parece não se sustentar.
Outro problema para a TSP é que foram detectadas circunstâncias em que a seleção age contra a homogeneidade genética. Por exemplo, se todos num formigueiro são 'iguais', uma mesma doença pode eliminar todos, então haveria vantagem na diferença genética, que é o contrário do parentesco, pois mais variedade significa mais chance de resistência a doenças.
Os autores também criticam os pressupostos da aptidão inclusiva como restritivos demais para a maioria dos casos em que se observa cooperação, e tentam demonstrar que usar a aptidão inclusiva é um desvio desnecessário que gera os mesmos resultados dos modelos que assumem seleção natural sobre aptidão simples. Ou seja, não é preciso complicar quando a teoria da seleção natural tradicional faz as mesmas previsões sem ser restritiva.
Mas nem tudo é crítica: Nowak, Tarnita e Wilson têm uma proposta alternativa para explicar a origem da eussocialidade:
Primeiro, a espécie deve formar grupos dentro de uma população, em casos como em localidades discretas atrativas para fazer ninhos ou procurar alimento, ou no caso de pais conviverem com filhotes, ou no caso de um rebanho seguir um líder; ou, ainda, grupos formados por acaso.
Depois, ocorre o acúmulo de características de predisposição à eussocialidade através da seleção natural comum. Por exemplo, certas abelhas solitárias dos gêneros Ceratina e Lasioglossum cooperam na busca de alimento, construção de túneis e armazenamento de recursos quando são forçadas experimentalmente à convivência. Outra predisposição seria a vigilância de ninhos como ocorre em várias vespas. Nesta etapa é invocada a "radiação adaptativa" - algumas espécies, uma minoria, serão propensas à eussocialidade (o que explica por que a eussocialidade é rara no número de espécies apesar dos organismos eussociais ocuparem uma fração enorme da biomassa do planeta).
Então, mutações e recombinação devem fixar genes ligados ao comportamento eussocial, servindo como gatilho para estabelecer as antigas predisposições em características estáveis de eussocialidade. Estes genes são desconhecidos, mas na formiga Solenopsis invicta, por exemplo, variantes do gene Gp-9 estão associadas à perda da capacidade de reconhecimento de operárias de outros formigueiros - o que explica as supercolônias dessa formiga no sul dos EUA.
Na quarta fase, a seleção natural favorece e estabiliza características emergentes de interação entre os organismos eussociais. Esta fase gera colônias que se comportam como superorganismos - as rainhas são como os gametas, e as operárias são como as células somáticas (as células de um organismo que executam funções variadas mas não passam seus genes adiante diretamente como fazem os gametas).
Finalmente, estes superorganismos podem sofrer seleção natural entre si (como acontece com os organismos comuns) - daí vem o refinamento das castas.
Mas por que, então, tanto parentesco entre organismos que cooperam, como nas formigas? O parentesco seria não a causa, mas a consequência da eussocialidade.
Se o modelo de Nowak, Tarnita e Wilson estiver correto, para que possamos compreender a evolução do comportamento social devemos adotar a noção de que há vários níveis para a seleção natural - dos genes aos superorganismos. Por isso, tratar apenas da fração de compartilhamento de genes, ou seja, parentesco, seria insuficiente.
E quanto aos humanos?
Este modelo, como ficou claro, deve ser ajustado para falar de humanos. A primeira coisa a ser feita é perceber que não somos eussociais em sentido estrito - nossas sociedades não lembram superorganismos, e apesar da tentativa histórica da Índia, não nos dividimos em castas naturalmente.
Outra diferença é que, diferentemente dos insetos, nós temos cultura. Nossos hábitos culturais são há milênios fonte de pressões seletivas sobre nossos genes, e a cultura co-evolui com eles. Ter cultura pressupõe nosso aparato cerebral; e a complexidade das nossas culturas está diretamente relacionada a capacidades mentais como a "teoria da mente" e a empatia - e esta tem tudo a ver cooperação, principalmente quando se observa coisas como adoção de crianças por casais gays não aparentados a elas!

Não foi por acaso que escolhi o exemplo dos casais gays tentando adotar crianças. Já se tentou explicar a origem da homossexualidade pela teoria da seleção de parentesco - o comportamento homossexual seria selecionado positivamente em sua origem porque os gays cuidam de seus sobrinhos (aptidão inclusiva). Os psicólogos evolutivos Paul Vasey e Doug VanderLaan, da Universidade de Lethbridge, Canadá, observaram no povo da ilha de Samoa, no Pacífico, que os homens gays de lá cuidam mais de sobrinhos do que os homens heterossexuais e as mulheres. Em Samoa, a cultura local vê os homens gays como um terceiro sexo que chamam de fa'afafine. Os pesquisadores explicam o comportamento dedicado dos tios fa'afafine em termos da seleção positiva de genes ligados à atração sexual por pessoas do mesmo sexo, mas não sabem até que ponto isso também pode ser explicado pela baixa homofobia da cultura samoana.
Se as críticas do grupo de E. O. Wilson à teoria da seleção de parentesco procedem, as conclusões de Vasey e VanderLaan são mais fracas do que se pensava. Em caso contrário, a TSP poderá continuar sendo um recurso para estudar a origem de comportamentos humanos como a homossexualidade e o altruísmo. Só o tempo dirá.
Enquanto isso, outros modelos explicativos alternativos à TSP são aventados. Relatando em 2008 à revista científica PLoS ONE, os italianos Andrea Camperio Ciani e Giovanni Zanzotto, da Universidade de Padova, e Paolo Cermelli, da Universidade de Torino, explicam a origem da homossexualidade por um modelo baseado na ideia de seleção sexualmente antagônica, na qual fatores genéticos espalham-se na população ao conferir vantagem reprodutiva para um sexo enquanto causa desvantagens ao outro. Neste caso, vantagem ao sexo feminino - a homossexualidade masculina estaria associada a uma maior fecundidade feminina - mães com filhos gays teriam mais filhos que outras mães. São resultados que também necessitam de aprofundamento.
Voltando à adoção, que é o comportamento de altruísmo social que estamos tentando explicar, vale lembrar que, ainda que a cultura seja um complicador na análise das origens da socialidade humana, não se pode negar a participação dos genes - logo, da evolução biológica - em sua origem. Richard Ebstein e seus colaboradores lembram, na revista Neuron, que há uma clara associação entre o comportamento social humano e genes que codificam receptores de neurotransmissores como oxitocina, vasopressina, serotonina e dopamina. Mutações nesses genes foram ligadas a transtornos de comportamento como o autismo (em que a "teoria da mente" e a empatia mostram-se deficientes) e a esquizofrenia.
Qualquer que seja a explicação evolutiva para a origem da nossa sociabilidade, expressada em comportamentos altruístas como a adoção de crianças por casais gays, uma coisa já sabemos: não é recente nem exclusiva.

Christophe Boesch e colegas, na revista PLoS One, relataram 18 casos de chimpanzés adotando jovens órfãos. Sendo os chimpanzés nossos parentes vivos mais próximos, parece que o altruísmo para com indivíduos não aparentados em nossos grupos é algo com mais de 7 milhões de anos, tempo em que nossas linhagens se separaram.
Fazer o bem sem olhar a quem é coisa nossa, dos primatas. Já as formigas, continuam sendo exemplo de eficiência, mas não de individualidade.
Imagens:
Formiga: JEAN-PHILIPPE VARIN / JACANA / SCIENCE PHOTO LIBRARY
Esquema sobre aptidão: modificado de Nowak et al. 2010.
E. O. Wilson: Public Library of Science.
Referências:
Nowak MA, Tarnita CE, & Wilson EO (2010). The evolution of eusociality. Nature, 466 (7310), 1057-62 PMID: 20740005
Boesch C, Bolé C, Eckhardt N, & Boesch H (2010). Altruism in forest chimpanzees: the case of adoption. PloS one, 5 (1) PMID: 20111704
Laland, K., Odling-Smee, J., & Myles, S. (2010). How culture shaped the human genome: bringing genetics and the human sciences together Nature Reviews Genetics, 11 (2), 137-148 DOI: 10.1038/nrg2734
Vasey PL, & VanderLaan DP (2010). An adaptive cognitive dissociation between willingness to help kin and nonkin in Samoan Fa'afafine. Psychological science : a journal of the American Psychological Society / APS, 21 (2), 292-7 PMID: 20424059
Camperio Ciani A, Cermelli P, & Zanzotto G (2008). Sexually antagonistic selection in human male homosexuality. PloS one, 3 (6) PMID: 18560521
Ebstein, R., Israel, S., Chew, S., Zhong, S., & Knafo, A. (2010). Genetics of Human Social Behavior Neuron, 65 (6), 831-844 DOI: 10.1016/j.neuron.2010.02.020