
Lição da evo-devo: Existe mais de um jeito de se fazer bicos e vários bicos que são feitos da mesma maneira

Compreender a evolução morfológica dos animais e plantas depende de entendermos mas a fundo a ontogenia dos organismos multicelulares, isto é, os padrões de mudança pré- e pós-natais que vão desde a formação do zigoto até a idade adulta, ou mesmo a morte, desses organismos. Pois é através de mudanças nestas etapas que novas formas e estruturas emergem ao longo da evolução a partir de modificações nas estruturas e formas anteriores por meio de alterações nos sistemas de controle genético-desenvolvimentais que estão a eles relacionados.
[Ao lado] Os tentilhões touro maiores das Antilhas (Loxigilla violacea) usam seus bicos profundos e largos para esmagar sementes e frutos duros. (Foto de José M. Pantaleón)
Certas criaturas mostram-se ótimos sistemas de estudo para investigar estas mudanças nos padrões, e nos sistemas de controle que estão por trás deles, uma vez que em um único grupo de seres vivos podem ser encontradas várias morfologias distintas que evoluíram a partir de um único ancestral comum nas diversas linhagens derivadas dele. Os bicos das aves são um desses sistemas de estudo óbvios, uma vez que é notório que a diversidade de forma e função em bicos de aves especializadas é abundante.
Foi Charles Darwin, ao coletar e estudar os tentilhões das ilhas Galápagos, correlacionando a morfologia (isto é, as formas) dos bicos das várias espécies (como John Gould mostraria mais tarde) aos tipos de distintos de sementes que cada espécie se especializara, que deu o primeiro impulso nesta linha de investigação que tem continuado até hoje cada vez de maneira mais completa e profunda.
Há alguns anos, em 2010 [1], por exemplo, um grupo de biólogos e biomatemáticos da Universidade de Harvard já haviam mostraram que os tentilhões de Darwin (Geospiza spp), na verdade, mesmo exibindo bicos morfologicamente bem distintos e adequados ao seus hábitos dietários, todos compartilharam as mesmas vias de desenvolvimento, isto é, usavam as mesmas redes de produtos gênicos que eram responsáveis pelo controle do tamanho e da curvatura dos seus bicos, dando origem 14 tipos de bicos bem diferentes uns dos outros, através de modificações nos padrões especiais e temporais de interação dos produtos dessas redes gênicas. Esta

impressionante variação de formas é gerada por alterações nas vias de sinalização que regulam o crescimento das duas componentes do esqueleto do bico: a cartilagem pré-nasal (PNC) e o premaxilar (PMX) [1].
Neste trabalho anterior publicado no periódico PNAS, o time de cientistas e matemáticos puderam mostrar que todas as formas dos bico das várias espécies de ‘tentilhões de chão’ (gênero Geospiza) estão relacionados por transformações de escala e a mesma relação vale também para todas as formas bico dos tentilhões das árvore, cocos e toutinegras (três gêneros distintos). As formas dos dos bico dentro de cada um desses grupos diferem apenas por suas escalas, tais como o comprimento e profundidade, que são geneticamente controlados pelas proteínas, ’proteína morfogenética óssea 4’ (Bmp4) e ’calmodulina’ (CaM) [1].
Os pesquisadores medindo a expressão da Bmp4 no primórdio bico das espécies do género Geospiza, conseguiram mapear a morfologia de maneira quantitativa as relações entre o bico e os níveis de expressão desta proteína. O modelo quantitativo consegue explicar as variações morfológicas dos bicos dentro dos tentilhões de Darwin por meio da extensão das transformações de escala para uma classe de transformações mais amplas (as do grupo afim) a partir da inclusão de transformações de cisalhamento [1].
Mais recentemente, os mesmo grupo de cientistas, expandindo esta linha de investigação, estudando um grupo de aves menos estreitamente relacionado, os tentilhões touros (’bullfinches’) das ilhas do Caribe, descobriram algo também muito interessante: os sinais moleculares que produzem essas formas de bico mostram uma variação ainda maior do que era possível desconfiar a partir de suas morfologias [2]. Os pesquisadores mostraram este grupo de pássaros do caribe, Loxigilla spp., evoluíram de maneira independente bicos com uma forma nova, diferente das exibidas pelas espécies do Geospiza, com as quais são estreitamente aparentados, mas também diferentes uns dos outros, distinguindo apenas em questão de escala. O surpreendente é que embora as formas gerais sejam as mesmas, as vias de sinalização e os padrões dos tecidos que formam os bicos das aves do gênero Loxigilla são diferentes nas três espécies [2].
Por exemplo, em Loxigilla noctis, assim como nas espécies de Geospiza, a cartilagem pré-nasal (PNC) desenvolve-se primeiro lugar, moldado através da sinalização por meio da Bmp4 e CaM, o que é seguido pelo desenvolvimento da região pré-maxilar (PMX) que é regulada pelos proteínas dos sistemas de sinalização que envolvem a TGFβIIr (veja mais sobre a via do TGF), β-catenina, e Dkk3. Porém, isos não é assim durante a morfogênese do bico da espécie Loxigilla violacea e nem em Loxigilla portoricensis em que a morfologia geral do bico é quase exclusivamente produzida a partir de modificações no PMX por meio de um mecanismo em que Ihh e Bmp4 interagem de maneira sinergistica para promover a expansão do tecido ósseo [2,3].
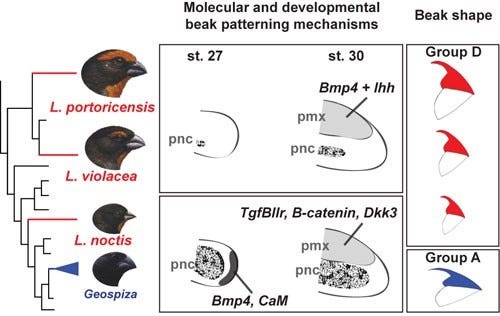
Acima podemos observar uma comparação entre os programas de desenvolvimento e as respectivas formas de bico que revela um elevado grau de flexibilidade nesta relação. Os tentilhões de Darwin (Geospiza, canto inferior esquerdo) e o tentilhões touro (“Bullfinch”) menores das Antilhas empregam tecidos semelhantes e basicamente os mesmos genes para estabelecer formas bico muito distintas, enquanto as espécies maiores desses tentilhões touro do Caribe usam uma combinação diferente de moléculas de sinalização para construir um bico que é basicamente idêntico das formas menores do mesmo gênero. (Imagem cortesia de Ricardo Mallarino.)
Os cientistas concluem que tais resultados demonstram uma elevada flexibilidade na relação entre morfologia e causas genético-desenvolvimentais subjacentes, já que vias de sinalização desenvolvimental podem dar origem a formas idênticas, e vias muito similares podem dar origem a formas bem diferentes, a partir de algumas ligeiras modificações. Assim, não só bicos muito diferentes podem originar-se a partir da mesma via de desenvolvimento, como é o caso dos tentilhões de Darwin, como morfologias de bicos muito semelhantes podem ser produzidas por vias de desenvolvimento bem distintas, como é o caso de Loxigilla [2,3],apontando para a evolução fenotípica convergente.
Este tipo de evidências são de extrema importância pois mostram claramente tanto que padrões divergentes podem ser conseguidos ao longo da evolução de uma linhagem com pequenas modificações de um sistema molecular e tecidual subjacente comum, dando origem a diferenças morfológicassubstanciais, como fenótipos muito similares podem emergir a partir de combinações gênicas diferentes e por alterações morfogenéticas teciduais distintas, o que deixa bem claro que não existe a necessidade que os mesmos tipos de modificações moleculares ocorram na mesma ordem e de forma idêntica, para que muitos fenótipos sejam 'alcançados’ ao longo da evolução de um grupo, existindo muitas vezes várias alternativas de rotas evolutivas, multiplicando a probabilidade de que certas morfologias evoluam.
__________________________________________________
Literatura Citada e Recomendada:
Campàs O, Mallarino R, Herrel A, Abzhanov A, Brenner MP. Scaling and shear transformations capture beak shape variation in Darwin’s finches. Proc Natl Acad Sci U S A. 2010 Feb 23;107(8):3356-60. Epub 2010 Feb 16. doi: 10.1073/pnas.0911575107.
Mallarino, R., Campàs, O., Fritz, J. A,. Burns, K. J,Weeks, O. G., Brenner, M. P. Abzhanov, A. Closely related bird species demonstrate flexibility between beak morphology and underlying developmental programs. Proceedings of the National Academy of Sciences, 2012; DOI: 10.1073/pnas.1206205109
Perry, Caroline In birds’ development, researchers find diversity by the peck, Press Release www.seas.harvard.edu September 24, 2012 [Link]
———————————————-
Créditos das Figuras:
Modelo da BMP-4 via Swissmodel usando como molde a BMP2 [PDB: 2h62 Chain: D]
CaM via PDB - Dutta, Shuchismita and Goodsell, David Calmodulin Molecule of the Month August 2003 doi: 10.2210/rcsb_pdb/mom_2003_8
Grande abraço,
Rodrigo