
Penas, para que te quero?
Já faz tempo que a imagem icônica dos dinossauros não é aquela típica dos desenhos e filmes feitos até meados dos anos 60 e 70. Lagartos agigantados e lerdos que arrastavam sua caudas de maneira pesada e desajeitada. Mesmo as representações que já mostram os dinossauros como animais ágeis e ativos, porém ainda com escamas grossas e reptilianas, fomentadas por filmes bem mais recentes (pós revolução do CGI), como Jurassic Park e sua seqüências, já começam a dar a impressão de desatualização. Frente aos belíssimos fósseis escavados e analisados nas ultimas décadas, em um espaço cada vez menor de tempo, nossa visão sobre estes incríveis animais vem se alterando
ainda mais, bem como nossa apreciação da complexidade e diversidade dessas criaturas.

Uma dessas mudanças de perspectiva é a crescente constatação de que os dinossauros não se extinguiram. As aves modernas não são, somente, os animais viventes mais aparentados com os dinossauros, elas próprias são dinossauros. Este grupo de vertebrados são os remanescentes da grande extinção do final do período cretáceo,
cerca de 65 milhões de anos atrás. Que extinguiu os demais dinossauros. Esta proposta ainda encontra alguma resistência de certos autores no meio cadêmico. Porém esta idéia parece cada vez mais difícil de ser negada. As aves, muito provavelmente, se originaram a partir de nossauros terópodes do caldo coelurossáuria, mais especificamente de um subgrupo chamado maniraptora, o mesmo grupo taxonômico ao qual pertencem os famosos velocirraptores.
As tentativas de achar furos nas filogenias que colocam as aves como um subgrupo dos terópodes (buscando enfatizar as diferenças entre os dinossauros “não-avianos” e os dinossauros “avianos”) não tem surtido muito efeito sobre a comunidade científica. A grande maioria dos paleontólogos vêem as aves, simplesmente como dinossauros vivos. Nenhuma hipótese filogenética alternativa é oferecida pelos críticos do modelo vigente, apenas vagas alusões de que as aves teriam se originado muito mais cedo, antes mesmo da origem dos terópodes, de um grupo desconhecido de Arcossauros (grupo ao qual tanto os dinossauros como os crocodilianos pertencem). Porém, a falta de fósseis evidenciando uma origem mais antiga para as aves, associada a fraqueza dos argumentos (que visam “demonstrar” que evolução das aves a partir dos dinossauros é improvável) é que são os maiores problemas das críticas a idéia de que as aves são dinossauros.
A despeito dessa controvérsia remanescente, uma coisa parece inegável. As penas, uma das mais distintivas características das aves modernas, simplesmente não “surgiram para o vôo”. A utilização das penas para o vôo parece ser uma co-optação, desta estrutura para uma nova função, relativamente tardia do ponto de vista filogenético (veja o artigo de Eli Vieira neste mesmo blog). Diga-se de passagem, a co-optação funcional (ou exaptação) de uma estrutura, e utilização dessa mesmas estrutura para diferentes funções em linhagens descendentes, parece ser algo muito recorrente na evolução.
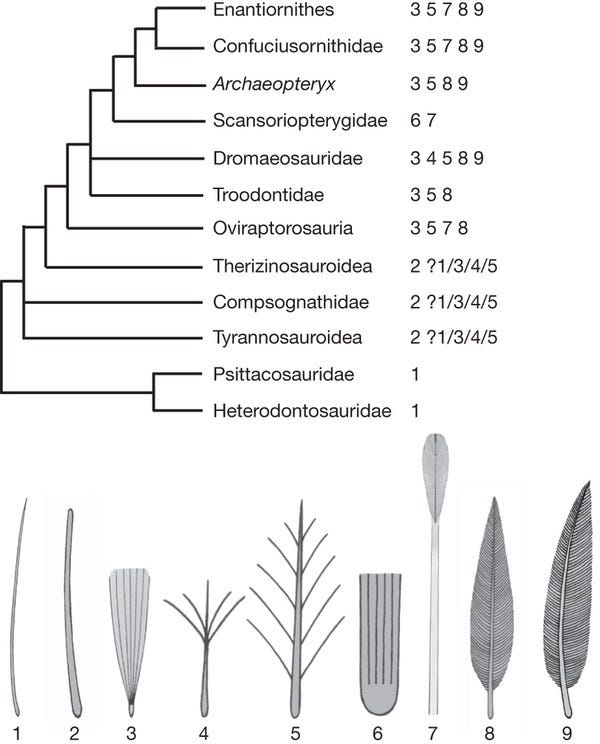
Nas últimas décadas evidências tem se acumulado mostrando que as penas já existiam em certos grupos de dinossauros, como os já mencionados grupos tetrápodes, mas muito possivelmente também, em outros grupos, como em certas linhagens de Ornithischia, uma das duas ordens que, junto com Saurischia (grupo formado pelos grande sarópodes e terópodes), formam o táxon Dinosauria.
Boa parte destas evidências vem de sítios fossilíferos localizados na China. Estas incríveis localidades tem revelado ao mundo muitos exemplares de dinossauros “não-avianos” com penas. Por isso, não é surpresa que, destes mesmos sítios, venha nossa mais recente janela para o passado longínquo. Desta vez os achados paleontológicos trazem pistas sobre o desenvolvimento ontogenético das penas nos dinossauros. Algo que pode motivar muito trabalho experimental e teórico na biologia evolutiva do desenvolvimento, a cada vez mais popular, “Evo-Devo”.

Em um artigo da Nature, Xu e colaboradores descrevem dois espécimes, STM4-1 e STM22-6, de dinossauros terópodes do cretáceo inferior, encontrados a oeste da província de Liaoning na formação de Yixian, China. O fato mas peculiar sobre estes dois espécimens é que ambos parecem ser indivíduos juvenis porém em fases diferentes de desenvolvimento. Já é bastante raro encontrar espécimens juvenis bem preservados, sobretudo com estruturas delicadas como penas, mas é ainda mais raro encontrar espécimens que ilustrem fases diferentes de desenvolvimento da mesma espécie.
Os dois espécimens possuem uma série de características derivadas partilhadas com outros maniraptores. Com crânios com uma região pré-orbital pequena; pré-maxila com uma grande região central, narinas externas dorsalmente posicionadas; uma grande fenestra mandibular e um longo processo retro-articular; além de um haste pubiana curvada anteriormente, ambos espécimes tem todas as indicações de serem pertencentes ao grupo de Oviraptorossauria, Similicaudipteryx, encontrados recentemente em escavações na mesma região.
STM4-1 possui características típicas de animais juvenis, com estrias longitudinais presentes nas superfícies dos ossos (muito possivelmente devido a calcificação incompleta) e arcos neurais não
fundidos na região central, com exceção da região da vértebra caudal. O espécimen também é muito menor do que adultos ou sub-adultos de outros fósseis de Oviraptorossauria, com um fêmur de 38 mm. Apesar de bem maior, com um fêmur de cerca de 140 mm, STM22-6 também parece ser um animal juvenil, já que os arcos neurais não estão ainda fundidos nas vértebras dorsal e caudal. Xu e
colaboradores sugerem portanto que STM4-1 e STM22-6 sejam respectivamente formas juvenis, iniciais e tardias de Similicaudipteryx.

As penas são estruturas epidérmicas que crescem a partir de folículos, como as escamas e os pelos em outros vertebrados, sendo formadas de uma substância chamada de queratina, a mesma que forma nossos cabelos e unhas. Existem vários tipos de penas, como as rêmiges e as retrizes, ambas chamadas “penas de vôo”, pelo menos nas aves modernas. As rêmiges cobrem as asas, possuindo contorno assimétrico, mais largas na sua parte interna; já as retrizes cobrem a cauda e tem contorno simétrico. Além delas, temos as tetrizes que são penas de cobertura e as plumas que são estruturas bastante delicadas que também revestem o corpo das aves. As penas podem ser subdivididas da seguinte forma: cálamo (a base oca) e ráquis (a haste central) que juntas formam o “eixo da pena” e as barbas e bárbulas que forma o “vexilo (ou lâmina) da pena”.
Os espécimens encontrados em Yixian tem uma combinação peculiar dessas estruturas e a disposição das penas e as características particulares delas em cada um dos espécimens sugerem que o desenvolvimento das penas ocorria de forma bastante diferente nesses animais em relação ao que acontece nos “dinossauros avianos”, ou seja, nas aves modernas. Xu e seus colegas sugerem que originalmente havia mais
diversidade e até flexibilidade desenvolvimental em relação as penas, mas que boa parte dela foi eliminada com a extinção das demais linhagens de terópodes.
As rêmiges são muito menores que as retrizes no espécimen STM4-1 do que em STM22-6, onde a diferença de tamanho é muito menos pronunciada. Esta outra diferença ontogenética em relação ao tamanho relativo das penas pode sugerir que as rêmiges desenvolviam-se em uma taxa desigual nos Similicaudipteryx, talvez “possivelmente refletindo o aumento no papel funcional das rêmiges em indivíduos se aproximando da fase adulta”.
No artigo, Xu, chama mais atenção para o contraste entre as rêmiges e retrizes do espécimen juvenil mais “crescido”, STM22-6, para as suas contrapartidas em STM4-1, o espécimen mais jovem, no qual as regiões proximais, ou seja, próximas a base, assumem a forma de laço e as regiões distais formando pontas “penosas”. Isto é algo que não parece ocorrer nas aves modernas que sofrem poucas mudanças após a primeira geração de penas. As rêmiges e retrizes em STM4-1 (aparentemente não podendo ser caracterizadas com primeira geração de penas) e STM22-6 são bastante diferentes umas das outras, sugerindo, segundo Xu, que mudanças morfológicas significantes dos tipos de penas ocorreriam mesmo na fase logo após a eclosão dos ovos. Algo que não é conhecido em nenhuma ave moderna.
A presença desses dois morfo-tipos (STM22-6 e STM4-1) indica que um processo de “muda” de penas, como nas aves modernas, já ocorria em Similicaudipteryx como parte do seu desenvolvimento ontogenético. A possibilidade alternativa de que essas diferenças representem apenas mudanças em uma única geração de penas parece muito menos provável, principalmente ao comparamos os tamanhos dos espécimens, julgando-os pelas diferenças do comprimento do fêmur, como o de STM4-1 sendo menos de 30% do comprimento do fêmur de STM22-6. Os autores vão mais além e sugerem que outros estágios deveriam existir além daqueles inferidos a partir dois morfo-tipos. Pelo menos mais um estágio de primeira geração de penas, e quem sabe estágios intermediários entre STM4-1 e STM22-6.
Penas “penáceas” (não plumosas) na forma de laços em sua porção distal (PRPFs) semelhantes as encontradas no espécimen STM4-1 também estão presentes em confuciusornitideos e outras aves ditas “primitivas”. Este tipo de pena parece ser amplamente distribuído tanto em grupos “basais” de aves como em “dinossauros não-avianos” maniraptores.
Três tipos de genes, Bone Morphognetic Protein (BMP), noggin e sonic hedgehog (SHH) desempenham um papel essencial na formação da raque e das barbas e do balanço entre estes elementos nas penas das aves modernas. Por este motivo, Xu e seus colaboradores, especulam que
alterações nos padrões de ativação dos genes relacionados com a especificação das barbas e da apoptose (morte celular programada) das regiões inter-barbas, provocados por genes “apoptóticos
inter-barbas”, poderiam gerar os morfo-tipos representados pelos dois espécimens estudados. Uma possibilidade envolveria a supressão do gene SHH que poderia formar vexilos contínuos que seriam “basicamente laminas de queratina não divididas em barbas”. Outra possibilidade é que este padrão poderia ser gerado pela expressão incompleta dos genes especificadores das barbas e apoptóticos inter-barbas. Estes genes são expressos muito cedo no desenvolvimento das gerações de
penas das aves modernas, mas ao comparar os padrões de desenvolvimento sugeridos pela análise dos dois morfo-tipos, os autores sugerem que a ativação, em Similicaudipterix, aconteceria de forma mais tardia e espacialmente incompleta produzindo a morfologia característica do morfo-tipo mais jovem, SMT4-1, apenas parcialmente coberto por penas penáceas. Este padrão então se modificaria nas fases mais tardias e, provavelmente, na fase adulta, onde a expressão dos genes especificadores das barbas e apoptóticos inter-barbas seriam expressos de forma equivalente ao que acontece nas aves modernas.
A expressão limitada do mesmo subconjunto de genes desenvolvimentais também poderia gerar outro tipo de penas do tipo elongadas amplas e filamentosas (EBFF) em outros tipos dinossauros terópodes encontrados recentemente. Estes achados, tomados em um contexto maior, levaram Xu e seus colaboradores a propor que as etapas de desenvolvimento inferidas a partir dos dois espécimens de Similicaudipterix e das recentes descobertas de dinossauros “não-avianos” basais com penas do tipo PRPF e EBFF, eram produzidas por um sistema genético semelhante ao atual, mas no qual a expressão de certos genes como especificadores das barbas e apoptóticos inter-brabas eram
incompletas ou mesmo completamente ausente. Além disso, apesar dos achados relatados no artigo e outros dados recentes serem consistentes com o modelo padrão de evolução das penas (barbas
→ ráquis → vexilo) a origem das penas parece ter sido mais complicada, inclusive com uma
maior flexibilidade e diversidade que simplesmente se perdeu durante a evolução e não está mais representada nas espécies de terópodes modernos, as aves. A filogenia proposta pelos autores aponta para esta diversidade original e sugere que os tipos 1,2,7 tenham se perdido com a extinção dos dinossauros não avianos.
Os estudos do grupo de Xu e outras novas descobertas, e análises de fósseis de dinossauros do período cretáceo, nos ajudam a
compreender como a evolução “molda” os fenótipos através de
alterações ontogenéticas, revelando nuances que a pouco tempo não eram nem imaginadas. Este trabalhos jogam um pouco mais de luz sobre o problema do desenvolvimento de novas características e, ao mesmo tempo, permitem-nos apreciarmos melhor o fato que a diversidade atual é só uma fração daquilo que já existiu.
Referências:
Xu, X., Zheng, X., & You, H. (2010). Exceptional dinosaur fossils show ontogenetic development of early feathers Nature, 464 (7293), 1338-1341 DOI: 10.1038/nature08965
Vieira, E. 2008. Penas: marcas indeléveis da evolução das aves.; Evolucionismo.ning.com, acesso em 05/05/2010.