
Sobre revistas, criacionismo e portais de imprensa. Parte II
Continuo com as considerações sobre o texto de Michelson Borges no Observatório da Imprensa, iniciadas na parte I ("Sobre revistas, criacionismo e portais de imprensa Parte I"), mas agora detendo-me mais no que está implícito em algumas dos questionamentos e críticas sobre a adequação científica da evolução dos seres vivos.
--------------------------------------------
Seria a biologia evolutiva uma ciência de segunda?
Existem, porém, outros argumentos implícitos nos questionamentos de Borges que merecem um olhar mais atento. O primeiro destes argumentos crípticos seria o de que apenas as “ciências experimentais” seriam válidas. O segundo seria o de que a macroevolução não passa pelo crivo dessas “ciências experimentais”. Estas sugestões originam-se de uma perspectiva simplista e equivocada sobre o que é a empreitada científica; uma visão que não tem qualquer respaldado na análise da própria prática científica feitas pelos filósofos, historiadores das ciências ou mesmo pelos cientistas mais reflexivos.

Uma das características principais das diversas ciências é a importância atribuída aos testes de hipóteses, modelos e teorias com base em dados empíricos coletados de forma metódica e analisados de maneira sistemática e crítica, sejam estes dados provenientes de estudos experimentais controlados ou observações minuciosas. Na realidade, muitos modelos e teorias de alto nível dependem de ambos os tipos de testes, experimentais e observacionais. O argumento criacionista implícito nas questões de Borges tem como objetivo ridicularizar e enfraquecer o papel das evidências observacionais na investigação científica, mas como vamos ver isso é um terrível equívoco.
Embora, como fazem alguns pesquisadores como é o caso da filósofa Caroll E. Clealand [50, 51, 52, 53, 54], possamos pensar em dois modos principais de se fazer ciência - o modo experimental, típico de campos da pesquisa como a física, a química e a biologia molecular, e o modo histórico, típico da astronomia, geologia, paleontologia etc – daí não podemos concluir que as ciências ditas históricas sejam epistemicamente inferiores as ditas experimentais, como a própria Clealand deixa bem claro [53, 54].
“As hipóteses alvo da maioria dos cientistas históricos são determinados eventos em grande escala do passado, por exemplo, um evento de extinção em massa específico, como o do fim do Cretáceo, ao contrário de extinções em massa em geral. Tais acontecimentos não podem ser testada diretamente em um cenário laboratorial, porque eles são irrepetíveis e, além disso, em muito grande escala para serem replicados artificialmente em laboratório. Dada esta diferença de enfoque científico não é de surpreender que as práticas dos cientistas históricos diferem de maneira significativa daqueles da ciência experimental clássico. Pesquisa histórica prototípica exibe um padrão distintivo de raciocínio probatório caracterizada por duas fases inter-relacionados: (1) a proliferação de vários hipóteses alternativas concorrentes, para explicar um corpo intrigante de traços (efeitos atuais de causas passadas), encontradas no trabalho de campo, (2) a busca por uma "arma fumegante" para discriminar entre elas. A arma fumegante discrimina entre hipóteses rivais históricas, revelando que uma (ou mais) fornecem uma explicação melhor para o corpo total de evidências disponíveis do que os outras.” [aqui]
Uma das principais diferenças entre as ciências históricas e experimentais é o papel da experimentação em ambientes controlados no teste de hipóteses. Porém, nas ciências históricas existem formas alternativas e muito robustas de testar hipóteses, o que inclui fazer predições específicas a serem confirmadas pela observação, bem como a utilização de múltiplas linhas de evidência “consilientes” e complementares [Veja este texto do NCSE sobre a questão da confusão criacionista, típica, opondo, de maneira simplória, “Ciências Experimentais vs Ciências Históricas”, fazendo isso mais comumente sobre a alcunha de “Ciências Históricas vs Ciências Operacionais” que não tem respaldo na historiografia moderna das ciências, por sinal].
Clealand argumenta que, embora os padrões de evidência possam ser diferentes, eles baseiam-se em uma assimetria objetiva e profundamente disseminada na natureza entre passado e futuro [53, 54]. Esta assimetria vem da sobredeterminação causal de eventos passados por eventos localizados presentes, que é o que explica a prática das ciências históricas, em contraste com subdeterminação causal de eventos futuros por eventos localizados presentes que, de modo complementar, explica as “ciências experimentais”, mais especificamente a necessidade delas empregarem controles estritos e manipulação diferencial das variáveis, uma vez que as condições testadas em um experimento são apenas causas parciais do que acontece posteriormente. Desta maneira, há sempre uma necessidade de explicitar e controlar outros fatores causais adicionais, caso contrário abrem-se muitas chances para falsos positivos e falsos negativos, ou seja, confirmações e refutações espúrias. Mas esse não é o caso das evidências de natureza histórica [53, 54].
“As diferenças entre os cientistas naturais, históricos e experimentais clássicos em metodologia são subscritas por um assimetria temporal pervasiva de causalidade bem conhecida dos físicos (Cleland, 2002). Esta assimetria de sobredeterminação (como foi denominada pelos filósofos) consiste no fato de que os eventos mais locais (em sentido amplo, de modo a incluir materiais e estruturais) 'sobredeterminam' suas causas passadas (porque o último normalmente deixa efeitos extensos e variados) e subdeterminam os seus efeitos futuros (porque raramente constituem a causa total de um efeito); colocando de maneira simples, o presente contém registros do passado, mas não há registros de futuro. Cientistas históricos exploram a sobredeterminação dos eventos passados por seus efeitos localizados de hoje, buscando vestígios denunciadores (isto é, uma 'arma fumegante') de eventos passados hipotéticos, e porque a maioria dos eventos deixam muitos desses traços no ambiente, eles jamais pode descartar completamente encontrá-los; é isso que justifica a busca de uma causa comum, quando confrontados com um conjunto de traços intrigantes descobertos através do trabalho de campo (Cleland, 2011). Como ilustração considere uma erupção vulcânica explosiva. Seus efeitos incluem extensos depósitos de cinzas, restos piroclástico, massas de magma andesitico ou riolitico, e uma grande cratera. Apenas uma pequena parte deste material é necessária para inferir a ocorrência da erupção. Na verdade, qualquer um de um número enorme de extremamente pequenas subcoleções de efeitos será sificiente. Isso ajuda a explicar por que os geólogos podem confiantemente inferir a ocorrência de evento do passado remoto, como as maciças erupções formadoras de caldeiras, que ocorreu 2.1 milhões de anos no que é hoje o Parque Nacional de Yellowstone.”[aqui]
Em alguns casos é bem obvio por que estudos experimentais, em sentido tradicional, são inviáveis. Afinal, como poderíamos reproduzir em grande escala fenômenos como a deriva continental e a tectônica de placas, ou replicar em laboratório algumas das grandes transições morfológicas que demoraram milhões de anos e que dependeram de centenas ou milhares de eventos de especiação (cladogênese) e de evolução de dentro de cada linhagem? Em outros casos, mesmo a exigência de observação direta é uma demanda absurda para alguns desses fenômenos e processos já que ou, no caso dos eventos em nosso passado, precisaríamos de uma máquina do tempo para testemunhá-los por nós mesmos, ou, nos casos de processos estendidos no tempo, a observação direta torna-se inviável por que ela excede a duração da vida humana ou mesmo a da nossa sociedade. Porém, como Clealand explica, existem vários dados disponíveis hoje em dia e que são escrutináveis pela comunidade científica que nos permitem inferir com alto nível de confiança o que ocorreu no passado e estabelecer que tipo de processos devem ter sido responsáveis por eles, testando a plausibilidade e adequação empírica relativa das diferentes alternativas, inclusive de uma maneira bem quantitativa.
Na verdade, é exatamente isso que é feito, por exemplo, com os estudos sobre as grandes transições evolutivas como as ilustradas pelas séries de fósseis com estruturas em estados transicionais entre formas viventes e extintas. Estas evidências conjuntas, que envolvem estudos filogenéticos, de anatomia comparativa, de séries estratigráficas, podem ser resumidas por meio de um dispositivo didático de apresentação gráfica chamado de evograma, como os que estão disponíveis no site do museu de Paleontologia da Universidade da Califórnia, Berkeley, dedicado ao ensino de evolução.
Os evogramas disponíveis no site Understanding Evolution incluem exemplos da transição dos vertebrados aquáticos para o terrestres, dinossauros para aves, da evolução dos cetáceos a partir de mamíferos terrestres quadrúpedes, surgimento dos mamíferos a partir de seus ancestrais sinapsídeos, além da emergência dos seres humanos. Através destas representações gráficas podemos perceber como o estudo anatômico comparativo de certas estruturas por meio dos fósseis de espécies extintas e modernas nos permitem apreciar os padrões intrincados de similaridade, o que nos permite perceber o nível de derivação da estrutura, da mais ancestral à mais derivada. Isso fica muito mais claro, principalmente, quando sobrepomos estas comparações anatômicas em uma filogenia baseada em outras características, ou seja, que não sejam as características transicionais investigadas, e que também mostra também os tempos de ocorrência daqueles grupos de organismos ao longo de milhões de anos. Neste tipo de esquema estão condensadas as evidências empíricas, os argumentos e as estratégias de inferência contidas em centenas de trabalhos científicos.
Estes estudos são apoiados por evidências moleculares e embriológicas dos grupos viventes, como por exemplo fica claro nos trabalhos que mostram a similaridade na expressão gênica em patas e nadadeiras de vertebrados terrestres e aquáticos [55, 56, 57] ou nos estudos embriológicos de espécies de mamíferos aquáticos em que podemos perceber os primórdios de estruturas como as patas traseiras durante o desenvolvimento inicial dos indivíduos, mas que que regridem antes no nascimento dos animais ou mesmo das vibrissas no rostro de certos cetáceos [58]. Outro ponto importante é que hipóteses sobre os padrões mais amplos e sobre os mecanismos agindo na evolução em grandes escalas de tempo e do espaço podem ser eles também testados por dados quantitativos paleontológicos, o que vem tornando-se cada vez mais comum graças ao grande número de bancos de dados computadorizados onde estão armazenadas informações taxonômicas, geográficas e contextuais precisas sobre espécimens fósseis, bem como por causa da existência de ferramentas de análise estatísticas e modelagem computacional e matemática, além de critérios de avaliação de escolha de modelos cada vez mais poderosos. Muitos dos estudos mais modernos empregam um conjunto de técnicas estatísticas e computacionais bastante complexas e refinadas que incluem os métodos de análise filogenética e de estimação dos tempos de divergência usando análise Bayesiana, modelos de máxima verossimilhança e calibração dos relógios moleculares por meio do registro fóssil [veja esta resposta de nosso tumblr], combinando-os critérios rigorosos de seleção de modelos, como AIC e outros. Nestas abordagens quantitativas, os dados de diferentes fontes de informação podem ser incorporados e hipóteses específicas podem ser testadas em relação a suas probabilidades posteriores ou verossimilhança. Alguns dos exemplos modernos mais impressionantes destas abordagens são o estudo publicado na Nature por Douglas Theobald [59, 60, 61, 62] [Veja também “Ancestralidade comum universal: A evidência está nas proteínas [Tradução]”], testando a hipóteses da ancestralidade comum universal usando um conjunto de proteínas ubiquais nos três domínios biológicos, e, mais recentemente, o trabalho que estimou as taxas de evolução morfológica e genética durante a explosão cambriana [63].
A questão, entretanto, é mais complexa, pois, como a própria Clealand deixa claro [54], há sobreposição entre as ciências de modo geral em relação aos seus métodos de estudo. Por exemplo, não é incomum vermos historiadores e arqueólogos usando métodos experimentais que vão desde as técnicas de datação por meio do radiocarbono, avaliação dietária por meio do estudo dos isótopos encontrados nos ossos e dentes, até a reconstrução de artefatos antigos por meios das técnicas supostamente empregadas pelos seus criadores originais em condições controladas e usando os recursos e conhecimento disponíveis em cada época, ou seja, a arqueologia experimental. Isso nos leva ao segundo argumento implícito na critica de Borges que seria a ideia que a evolução não teria apoio experimental. Esta sugestão é ridícula, pois a biologia evolutiva, como enfatiza Massimo Pigliucci [64], encontra-se bem no meio desta divisão metodológica entre ciências experimentais e históricas, usando abundantemente ambos os padrões de evidência de maneira bastante disseminada.
Existe uma gama enorme de estudos experimentais [veja também o verbete da wikipédia, além da edição especial da revista Biology Letters sobre o assunto] [64, 65] feitos em moléculas como o RNA [66] organismos com ciclos de vida mais curtos, como vírus, bactérias, protistas, algas, fungos, invertebrados (como os estudos em moscas de fruta feitos a partir dos trabalhos pioneiros de Dobzhansky), vertebrados (como camundongos, peixes e lagartos) [67, 68, 69, 70, 71, 72, 73, 74] e plantas que incluem tanto a investigação de processos microevolutivos, como aqueles realizados na pesquisa de como fatores evolutivos como a seleção natural, deriva genética e mutações interagem ao longo das gerações em populações de laboratório e selvagens, como aqueles conduzidos na investigação dos processos de especiação [Por sinal não custa lembrar dos vários exemplos de especiação, portanto, de divergência de populações e formação de novas linhagens que os cientistas acumularam ao longo dos anos. Veja “instâncias observadas de especiação” e “mais instâncias observadas de especiação”], muitos dos quais envolvem submeter populações de uma mesma espécie a pressões distintas com objetivo de estudar a divergência genética e fenotípica, bem como os fatores e mecanismos responsáveis pelo isolamento reprodutivo, como estudo com híbridos obtidos entre espécies próximas de modo a revelar qual a base genética das diferenças fenotípicas e como elas surgiram ao longo do passado destas linhagens [65, 66], além de abordar questões como a origem e manutenção da reprodução sexual. Estes estudos envolvendo a especiação e a base genética de características fenotípicas distintivas entre espécies próximas já qualificam-se naquilo que os biólogos evolutivos consideram macroevolução, o que já serviria para desmentir as sugestões de Borges. Mas isso não é tudo.
Mesmo que não possamos recriar, em toda sua exuberância e complexidade, os eventos de transição que estudamos por meio das evidências fósseis, anatômicas, embriológicas e moleculares disponíveis hoje em dia, podemos, entretanto, realizar estudos experimentais cruciais para compreendermos estas transições e analisarmos os dados de maneira mais crítica e precisa. Por exemplo, muitos paleontólogos conduzem experimentos no que eles chamam de tafonomia experimental [75],estudando as etapas iniciais de fossilização e como os diferentes processos egológicos e biológicos interferem com as características destes vestígios; além disso existe uma área muito popular, atualmente, entre os biólogos evolutivos chamada de de EVO-DEVO (Biologia Evolutiva do Desenvolvimento) [76, 77, 78] em que abordagens da anatomia e embriologia comparativa, paleontologia, além do estudo da expressão gênica e da biologia molecular do desenvolvimento, são combinados com os modernos métodos de análise filogenéticos baseados em biomoléculas e morfologia. Neste campo, frequentemente, são empregados métodos experimentais envolvendo a manipulação controlada dos circuitos genéticos e das vias do desenvolvimento de certas estruturas, sistemas e órgãos, tidos pelos cientistas, como sendo responsáveis pelas mudanças ao longo da história evolutiva dos grupos que estão sendo investigados.
Este tipo de evidência experimental, mesmo indireta, é fundamental para os estudos sobre a macroevolução, já que nos permitem investigar mecanisticamente como alterações no material genético que, por exemplo, modificam o padrão espacial e temporal e a intensidade da expressão dos genes - e, desta maneira, alteram os padrões de interação entre células e tecidos ao longo do desenvolvimento ontogenético - podem ter dado origem a certas estruturas a partir das versões ancestrais das mesmas em outras espécies. Exemplos recentes deste tipo de estudo experimental incluem a indução de patas em peixes [79, 80] e de estruturas similares a nadadeiras em camundongos [81 82]. Veja também os textos "Viva Turing de novo, mais pistas sobre a evolução dos membros em vertebrados" e "Superexpressão do gene 13Hoxd: Mais pistas sobre a transição entre peixes e tetrápodes:"
Outro exemplo são os estudos experimentais mais relevantes a questão da origem da multicelularidade em que organismos modernos como leveduras, algas unicelulares e outros microrganismos são submetidos a regimes de seleção específicos que teriam sido equivalentes aos que, de acordo com os modelos e teorias evolutivas e evidências filogenéticas, teriam sido responsáveis pela evolução deste tipo de organismo [83, 84, 85, 86]. Veja também os textos "Evolução da multicelularidade em laboratório" e "Evolução da multicelularidade em laboratório II".
Nestas linhas de pesquisa científica é investigado, exatamente, aquilo que muitos criacionistas dizem não ser possível, ou seja, como surgem as inovações evolutivas. Por meio dos métodos e estratégias adotadas nesses ramos da pesquisa biológica podemos testar as hipóteses elaboradas com base em estudos filogenéticos e em análises comparativas da anatomia e embriologia dos organismos modernos e fósseis. O problema é que os criacionistas têm suas próprias definições sobre o que seria evolução e insistem em exigir 'evidências' que não fazem o menor sentido. Este é o caso, por exemplo, do 'crocopato' (de Kirk Cameron e Ray Confort) que reflete um equívoco mais geral sobre a evolução que é o de imaginar que as espécies transicionais seriam algum tipo de quimeras, compostas por partes de organismos modernos [veja por exemplo aqui e aqui]. Na mesma linha de equívocos podemos citar as demandas por parte de certos criacionistas de tipos de evidências incompatíveis com a realidade, como exigir observação direta das grandes transições, supondo que elas deveriam ocorrer em um único evento de divisão de linhagem equivalente a especiação, quando os cientistas estão cansados de explicar que tais transições não são eventos simples, mas fenômenos compostos de vários eventos de especiação e extinção de linhagens, aos quais, apenas em retrospecto, faz sentido tratarmos deles como eventos de formação de grupos taxonômicos mais amplos, como gêneros, famílias, ordens, classes e filos [14].
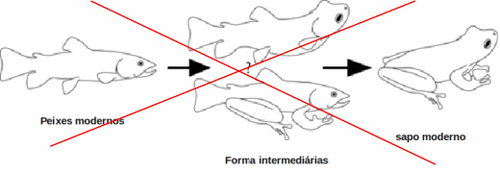
Ainda assim, existem fenômenos que ocorrem em algumas gerações e àqueles que testemunhamos através da história humana que, embora tecnicamente enquadrem-se dentro da microevolução, por não estarem associados a eventos de divisão de linhagens, mesmo assim, envolvem níveis de mudança fenotípica típicos de algumas das transformações que ocorrem cumulativamente em largas escalas de tempo. Por exemplo, a maioria dos tipos de alterações herdáveis que participam destas transições, envolvendo tamanho, proporções, coloração e número de partes de organismos multicelulares, já foram observadas em campo ou em laboratório [14]. O caso de espécies vegetais cultivadas como o milho são um exemplo ainda mais claro disso. Estes exemplos demostram como, em alguns milhares de anos, empregando a diversidade genética existente nas populações naturais e regimes de seleção direcional específicos foi possível promover grandes transformações morfológicas, como fica claro ao compararmos o milho atual ao seu parente o teosinte [87].
Ainda mais impressionantes são as diversas raças de cães obtidas por meio do cruzamento seletivo a partir de lobos, domesticados ao longo dos séculos. Além das alterações de tamanho, que deram origem aos Chihuahua e aos Dinamarqueses, existe uma gama enorme de variação em relação as proporções dos membros, da morfologia craniofacial, coloração e comportamento. Dois pesquisadores, Abby Grace Drake e Christian Peter Klingenberg [88], seguindo o trabalho anterior de Robert K. Wayne, feito em 1986 [89], resolveram quantificar as diferenças morfológicas através da estimação daquilo que os biólogos evolutivos chamam de disparidade. Por meio de técnicas de análise morfométricas, eles foram capazes de mostrar que a disparidade nos cães domésticos excede a exibida por qualquer outra espécie individual de canídeos. Na verdade, a disparidade dentro deste subgrupo é equivalente a disparidade de toda a ordem carnívora [88]. O fato destas alterações nos cães, no milho e em outros organismos domesticados não estarem ligadas a especiação - portanto, ao estabelecimento de linhagens distintas - é apenas um detalhe já que os tipos de modificação morfológica são equivalentes aos tipos de mudanças que estão por trás das diferenças entre grupos bem mais distantes de organismos, mas que ocorreram de forma cumulativa após muitos eventos de especiação, evolução dentro das linhagens e extinção. Desta maneira, estas modificações morfológicas que ocorrem em espaços de tempo mais curtos e que conseguimos rastrear em quanto sociedade é tão dramática que lançam luz ao estudo da macroevolução, nos permitindo estudá-la bem de perto e com muito mais precisão.
Por fim, existem outros métodos de investigar a macroevolução, como a utilização de simulações computacionais e modelagem matemática da evolução de características fenotípicas e genéticas ao longo da evolução tendo como base a moderna teoria da genética quantitativa e de populaççoes evolutiva e os métodos filogenéticos comparativos, em que diversos processos podem ser simulados tendo como base árvores filogenéticas de um dado grupo de organismos, de modo que possamos inferir os processos evolutivos envolvidos nem sua evolução [82, 83, 84]. O campo da Vida Artificial, especialmente de 'Evolução de Organismos Digitais' é outras maneira de investigar padrões e mecanismos evolutivos gerais, como podemos constatar através de trabalhos em plataformas como AVIDA, Polyword e Tierra [89, 90, 91, 92].
Todas essas abordagens complementam-se e reforçam-se umas as outras, nos permitindo explorar, não só a evolução dentro de populações, mas a evolução em largas escalas temporais e espacias, nas quais múltiplos eventos de especiação e extinção ocorrem, tendo como pano de fundo mudanças ecológicas e ambientais profundas. Claro. Como em qualquer campo científico, existem muitas dúvidas e questões em aberto; além do mais, os cientistas, compreendendo a falibilidade humana, estão preparados para novas evidências, teorias e argumentos que os façam refletir e, eventualmente, mudar de ideia, mesmo que não haja nada assim afigurando-se no horizonte. Esta postura crítica e a humildade epistêmica vinculada a ela são parte integrantes dos valores cognitivos compartilhados pela comunidade científica e são o que a diferencia de outras empreitadas não tão críticas e organizadas. Conclusões científicas como as sobre a idade do universo, do nosso sistema solar e planeta, bem como sobre a evolução dos seres vivos, são apoiadas por uma enorme gama de evidências que foram descobertas, avaliados e validadas por gerações de pesquisadores com as mais variadas posturas religiosas, filosóficas e políticas.
Incitar a não aceitação do fenômeno da evolução, apresentando-o como uma conclusão não científica e/ou como um mero jogo de pressão ideológica, e ignorar as conclusões da biologia evolutiva não é apenas errado. É profundamente perigoso. O estudo da evolução tem sim implicações práticas importantes [veja aqui, aqui, aqui] que vão desde aplicações da investigação investigação forense e criminal [veja aqui também] [97, 98] e chegam mesmo em áreas como a robótica e a computação. [Existe, inclusive, pelo menos um periódico dedicado a isso, o Evolutionary Applications.]
Em seguida cito apenas alguns exemplos mais bem conhecidos - que nem de longe esgotam as contribuições da biologia evolutiva e, portanto, da teoria evolutiva moderna - em outras questões mais aplicadas e de relevância social e política mais imediatas [99, 100, 101, 102, 103, 104, 105, 106, 107, 108, 109] :
1) Em nossa saúde, tanto ao nos permitir uma compreensão mais profunda da resistência a antibióticos, antivirais, antiparasitários e agentes outros quimioterápicos, bem como em nossa compreensão epidemiológica de várias doenças infecto-contagiosas (HIV, dengue, febre-amarela) e dos seus vetores (mosquitos, por exemplo) e mesmo dos mecanismos pelos quais as doenças emergem, muitos deles associados a forma como evoluímos;
2) Na produção de alimentos a biologia evolutiva permite uma compreensão melhor da evolução da resistência a pesticidas por parte de insetos e plantas;
3) Na conservação de espécies e ecossistemas e no manejo de recursos naturais (e mesmo em sua relação com a agricultura e pecuária) a biologia evolutiva nos ajuda a compreender como os mecanismos de adaptação são sensíveis às mudanças no seu meio ambiente, principalmente como as diretamente ligadas a atividade humana, como desmatamento, pesca excessiva ou a mudanças climáticas globais, além de nos dar um melhor entendimento dos processos de interação entre espécies (especialmente em relação ao problema das espécies invasoras e do impacto da remoção de predadores de topo nos ecossistemas) que mantém a estabilidade dos ecossistemas e sua dinâmica de curto, médio e longo prazo. Estes conhecimentos são essenciais a médio e longo prazo e ignorá-los poder ter consequências nefastas.
4) Na biotecnologia, como é o caso dos estudos de 'evolução dirigida' - o estudo da evolução e a aplicação de estratégias de seleção artificial em ambientes controlados abre espaço para a criação de novas moléculas, materiais e processos de produção. Outro exemplo é o da biorremediação. A biologia evolutiva, por meio da investigação dos processos e mecanismos de evolução e adaptação a ambientes extremos, possibilita a procura e seleção de espécies de microrganismos e plantas capazes de destoxificação de áreas contaminadas etc.
Tratar a evolução como uma mera questão de opinião não consiste somente em rejeitar uma cosmovisão específica, mas endossar um desprezo pelas evidências quando elas não dizem, exatamente, aquilo que queremos ouvir. É uma atitude de imaturidade epistêmica, mas que acima de tudo têm implicações sócias e políticas de médio e longo prazo que não deveriam poder ser ignoradas em uma sociedade como a nossa que depende tanto de informação de boa qualidade e dos conhecimentos científicos para resolver seus problemas e as mazelas sócias e ambientais. Compreender os problemas da oposição dos criacionistas às conclusões científicas é essencial, bem como compreender a natureza distorcida e mal informada de seus ataques ao ensino e divulgação da biologia evolutiva, de modo a denunciá-los e melhor combatê-los.
---------------------------------------
Referências:
Futuyma, D.J. Biologia Evolutiva. Funpec. Ribeirão Preto. 3ª Edição. 2009. 830 p
Freeman, S.; Herron, J C.. Análise Evolutiva. 4ª edição Porto Alegre: Artmed, 2009. 831 p.
Mayr, Ernst. O que é evolução. Rio de Janeiro: Roco, 1ª edição 2009. 342 p.
Ridley, Mark. Evolução 3ª edição Porto Alegre: Artmed, 2006. 752 p.
Gregory, T. Ryan Evolution as Fact, Theory, and Path Evolution: Education and Outreach January 2008, Volume 1, Issue 1, pp 46-52 DOI: 10.1007/s12052-007-0001-z
Hofmann, James R., Weber, Bruce H. The Fact of Evolution: Implications for Science Education Science & Education November 2003, Volume 12, Issue 8, pp 729-760, 2003 DOI: 10.1023/B:SCED.0000004556.80802.3a
Weber, B.H. "Fact, phenomenon, and theory in the Darwinian research tradition", Biological Theory 2(2), 168-178, 2007.
Wiles, J. R. (2010). Overwhelming scientific confidence in evolutionary theory and its centrality in science education--And the public disconnect. The Science Education Review, 9, 18-27. [Link]
Sesink Clee, P. & Gonder, M. K. (2012) Macroevolution: Examples from the Primate World. Nature Education Knowledge 3(12):2 -
Wilkins, John Macroevolution: Its Definition, Philosophy and History TalkOrigins Archives Version 2.1.3, Last Update: September 23, 2006. [Link]
Padian, K. 2010. How to Win the Evolution War: Teach Macroevolution! Evolution Education and Outreach, 3: 206-214. [Link]
Bowler, Peter J(Apr 2001) Evolutionary Ideas: The Modern Synthesis. In: eLS. John Wiley & Sons Ltd, Chichester. http://www.els.net [doi: 10.1038/npg.els.0001694]
Erwin DH. Macroevolution is more than repeated rounds of microevolution. Evol Dev. 2000 Mar-Apr;2(2):78-84. PubMed PMID: 11258393.
Theobald, Douglas L. "29+ Evidences for Macroevolution: The Scientific Case for Common Descent." The Talk.Origins Archive. Vers. 2.89. 2012. Web. 12 Mar. 2012.
Mayo, Oliver Charles Darwin as Experimenter: One of many Reasons to Remember him at 2009 Source: Transactions of the Royal Society of South Australia, Volume 133, Number 2, November 2009 , pp. 288-299(12)
Bowler, Peter J(Sep 2010) Evolution: History. In: eLS. John Wiley & Sons Ltd, Chichester. http://www.els.net [doi: 10.1002/9780470015902.a0001517.pub2]
Bowler, Peter J(Apr 2001) Evolutionary Ideas: The Eclipse of Darwinism. In: eLS. John Wiley & Sons Ltd, Chichester. http://www.els.net [doi: 10.1038/npg.els.0001693]
Takahata, Naoyuki(Dec 2007) Molecular Evolution: Neutral Theory. In: eLS. John Wiley & Sons Ltd, Chichester. http://www.els.net [doi: 10.1002/9780470015902.a0001800.pub2]
Ohta, Tomoko(Feb 2013) Molecular Evolution: Nearly Neutral Theory. In: eLS. John Wiley & Sons Ltd, Chichester. http://www.els.net [doi: 10.1002/9780470015902.a0001801.pub4]
Penny, David(Apr 2013) Molecular Evolution: Introduction. In: eLS. John Wiley & Sons Ltd, Chichester. http://www.els.net [doi: 10.1002/9780470015902.a0001701.pub3]
Sheldon, Peter R(Apr 2001) Punctuated Equilibrium and Phyletic Gradualism. In: eLS. John Wiley & Sons Ltd, Chichester. http://www.els.net [doi: 10.1038/npg.els.0001774]
Lieberman, Bruce S. and Eldredge, Niles (2008) Punctuated equilibria, Scholarpedia, 3(1):3806.[Link]
Damuth, John D(Apr 2001) Evolution: Tempo and Mode. In: eLS. John Wiley & Sons Ltd, Chichester. http://www.els.net [doi: 10.1038/npg.els.0001720]
Broom, Mark, and Cannings, Chris(Nov 2010) Evolutionary Game Theory. In: eLS. John Wiley & Sons Ltd, Chichester. http://www.els.net [doi: 10.1002/9780470015902.a0005457.pub2]
Alexander, J. McKenzie, "Evolutionary Game Theory", The Stanford Encyclopedia of Philosophy (Fall 2009 Edition), Edward N. Zalta (ed.).
Cowden, C. C. (2012) Game Theory, Evolutionary Stable Strategies and the Evolution of Biological Interactions. Nature Education Knowledge 3(10):6 [link]
Okasha, Samir, "Biological Altruism", The Stanford Encyclopedia of Philosophy (Fall 2013 Edition), Edward N. Zalta (ed.).
Crespi, Benard J(Apr 2001) Selection: Units and Levels. In: eLS. John Wiley & Sons Ltd, Chichester. http://www.els.net [doi: 10.1038/npg.els.0001712]
Lloyd, Elisabeth, "Units and Levels of Selection", The Stanford Encyclopedia of Philosophy (Winter 2012 Edition), Edward N. Zalta (ed.), URL = <http://plato.stanford.edu/archives/win2012/entries/selection-units/>;.
Arthur, Wallace(Nov 2011) Evolutionary Developmental Biology: Developmental Bias and Constraint. In: eLS. John Wiley & Sons Ltd, Chichester. http://www.els.net [doi: 10.1002/9780470015902.a0001066.pub3]
Futuyma, D. J. (2010), Evolutionary constraint and ecological consequences. Evolution, 64: 1865–1884. doi: 10.1111/j.1558-5646.2010.00960.x
Wagner A. Genotype networks shed light on evolutionary constraints. Trends Ecol Evol. 2011 Nov;26(11):577-84. doi: 10.1016/j.tree.2011.07.001.
Crow, James F(Mar 2008) Evolution: Views of. In: eLS. John Wiley & Sons Ltd, Chichester. http://www.els.net [doi: 10.1002/9780470015902.a0005104.pub2]
Gould, Stephen Jay The Structure of Evolutionary Theory The Belknap Press of Harvard University Press, 2002, 1433 pp
Carroll, R.L. A new evolutionary synthesis: do we need one? Trends in Ecology and Evolution, v. 15, 2000, pp. 205-206.
Carroll, S.B. (2008) Evo Devo and an Expanding Evolutionary Synthesis: A Genetic Theory of Morphological Evolution. Cell 134: 25-36.
Pigliucci, Massimo 2007. Do we need an extended evolutionary synthesis? Evolution 61 12, 2743–2749.
Müller GB. Evo-devo: extending the evolutionary synthesis. Nat Rev Genet. 2007 Dec;8(12):943-9. Review. PubMed PMID: 17984972.
Lewin, R., Foley, R.A., Principles of Human Evolution, 2nd Ed., Blackwell Science Ltd, 2004. 576 pg
Wood, B. Human Evolution: A Very Short Introduction.Oxford University Press, New York, 2005. 144 pg.
Perelman P, Johnson WE, Roos C, Seuánez HN, Horvath JE, et al. (2011) A Molecular Phylogeny of Living Primates. PLoS Genet 7(3): e1001342. doi:10.1371/journal.pgen.1001342
Finstermeier K, Zinner D, Brameier M, Meyer M, Kreuz E, et al. (2013) A Mitogenomic Phylogeny of Living Primates. PLoS ONE 8(7): e69504. doi:10.1371/journal.pone.0069504
Diogo, R. and B. Wood. "Soft-tissue anatomy of the primates: phylogenetic analyses based on the muscles of the head, neck, pectoral region and upper limb, with notes on the evolution of these muscles," Journal of Anatomy 219: 273–359, 2011.
Diogo, R., and B. Wood. Comparative Anatomy and Phylogeny of Primate Muscles and Human Evolution. Oxford, UK: Taylor and Francis. 2012
Mitchell, M. W. & Gonder, M. K. (2013) Primate speciation: A case study of African apes. Nature Education Knowledge 4(2):1 [Link]
Wood, B.A. and Richmond, B. ‘Human evolution: taxonomy and paleobiology.’ J. Anat., 97: 19-60. 2000. PDF
Kehrer-Sawatzki H, Cooper DN. Understanding the recent evolution of the human genome: insights from human-chimpanzee genome comparisons. Hum Mutat. 2007 Feb;28(2):99-130. Review. PubMed PMID: 17024666.
Pollard KS. What makes us human? Sci Am. 2009 May;300(5):44-9. PubMed PMID: 19438048.
Watanabe H, Hattori M. [Chimpanzee genome sequencing and comparative analysis with the human genome.] Tanpakushitsu Kakusan Koso. 2006 Feb;51(2):178-87. Review. Japanese. PubMed PMID: 16457209.
Yoko K, Atsushi T, Hideki N, Asao F. [Comparative studies on human and chimpanzee genomes]. Tanpakushitsu Kakusan Koso. 2005 Dec;50(16 Suppl):2072-7. Review. Japanese. PubMed PMID: 16411432.
Cleland, C. E. and Brindell, S. (forthcoming): Science and the Messy, Uncontrollable World of Nature, in Pigliucci, M and Boudry, M. (eds.), Philosophy of Pseudoscience: Reconsidering the Demarcation Problem (University of Chicago Press) (in press).
Cleland, C. E. (2011): Prediction and Explanation in Historical Natural Science, British Journal of Philosophy of Science 62, pp. 551-582. http://bjps.oxfordjournals.org/content/early/2011/02/25/bjps.axq024.full.pdf+html
Cleland, C. E. (2002): Methodological and Epistemic Differences Between Historical Science and Experimental Science, Philosophy of Science 69, pp. 474-496.
Cleland, C. E. (2001): Historical science, experimental science, and the scientific method, Geology 29, pp. 987-990.
Johanson Z, Joss J, Boisvert CA, Ericsson R, Sutija M, Ahlberg PE. Fish fingers: digit homologues in sarcopterygian fish fins. J Exp Zool B Mol Dev Evol. http://onlinelibrary.wiley.com/doi/10.1002/jez.b.21197/abstract2007
Freitas R, Zhang G, Cohn MJ (2007) Biphasic Hoxd Gene Expression in Shark Paired Fins Reveals an Ancient Origin of the Distal Limb Domain. PLoS ONE 2(8): e754. doi:10.1371/journal.pone.0000754 [Link]
Davis MC, Dahn RD, Shubin NH. An autopodial-like pattern of Hox expression in the fins of a basal actinopterygian fish. Nature. 2007 May 24;447(7143):473-6. PubMed PMID: 17522683. [Link]
Thewissen, J.G.M., Cooper, L.N., George, J.C., and Bajpai, S. 2009. From land to water: The origin of whales, dolphins, and porpoises. Evolution: Education & Outreach 2:272-288.DOI: 10.1007/s12052-009-0135-2 [Link]
Theobald DL "A formal test of the theory of universal common ancestry." Nature 465(7295):219-222. [pdf]
Theobald DL (2010) "Was the universal common ancestry proved? Reply" Nature 468(7326):E10. [pdf]
Steel M, Penny D. Origins of life: Common ancestry put to the test. Nature. 2010 May 13;465(7295):168-9. doi: 10.1038/465168a. PubMed PMID: 20463725. [pdf]
Theobald DL. On universal common ancestry, sequence similarity, and phylogenetic structure: the sins of P-values and the virtues of Bayesian evidence. Biol Direct. 2011 Nov 24;6(1):60. PubMed PMID: 22114984; PubMed Central PMCID: PMC3314578. [pdf]
Lee, Michael S.Y., Soubrier, Julien, Edgecombe, Gregory D. Rates of Phenotypic and Genomic Evolution during the Cambrian Explosion. Current Biology, 2013 DOI: 10.1016/j.cub.2013.07.055 [Link]
Pigliucci, M. 2013. The Nature of Evolutionary Biology: At the Borderlands Between Historical and Experimental Science. In: The Philosophy of Biology: A Companion for Educators, ed. by K. Kampourakis, Springer, pp. 87-100
Garland, T., Jr., and M. R. Rose, eds. 2009. Experimental evolution: concepts, methods, and applications of selection experiments. University of California Press, Berkeley, California.
Kawecki, Tadeusz J. et al. Experimental evolution Trends in Ecology & Evolution - 1 October 2012 (Vol. 27, Issue 10, pp. 547-560) DOI: 10.1016/j.tree.2012.06.001
Domingo-Calap P, Sanjuán R. Experimental evolution of RNA versus DNA viruses. Evolution. 2011 Oct;65(10):2987-94. doi: 10.1111/j.1558-5646.2011.01339.x.
Bennett, Albert F., Hughes, Bradley S. Microbial experimental evolution American Journal of Physiology - Regulatory, Integrative and Comparative PhysiologyPublished 1 July 2009Vol. 297no. R17-R25DOI: 10.1152/ajpregu.90562.2008 [Link]
Dunham MJ. Experimental evolution in yeast: a practical guide. Methods Enzymol. 2010;470:487-507. doi: 10.1016/S0076-6879(10)70019-7. [Link]
terHorst CP. Experimental evolution of protozoan traits in response to interspecific competition. J Evol Biol. 2011 Jan;24(1):36-46. doi: 10.1111/j.1420-9101.2010.02140.x.
Zeyl, C. (2006), Experimental evolution with yeast. FEMS Yeast Research, 6: 685–691. doi: 10.1111/j.1567-1364.2006.00061.x
Bell G. Experimental evolution of heterotrophy in a green alga. Evolution. 2013 Feb;67(2):468-76. doi: 10.1111/j.1558-5646.2012.01782.x.
Burke, Molly K., Rose, Michael R. Experimental evolution with Drosophila American Journal of Physiology - Regulatory, Integrative and Comparative PhysiologyPublished 1 June 2009Vol. 296no. R1847-R1854DOI: 10.1152/ajpregu.90551.2008 -[Link]
Burke, Molly K. et al. Genome-wide analysis of a long-term evolution experiment with Drosophila Nature, 467, 587-590, (30 September 2010) | doi: 10.1038/nature09352
Briggs, D.E.G. 1995 Experimental taphonomy. Palaios 10, 539-550.
Carrol, Sean B. Infinitas Formas de Grande Beleza: Como a evolução forjou a grande quantidade de criaturas que habitam o nosso planeta. 1ª edição Rio de Janeiro: Jorge Zahar Editor, 2003. 303 p.
Carrol, Sean B. From DNA to Diversity: Molecular Genetics and the Evolution of Animal Design, with Jennifer Grenier and Scott Weatherbee, Wiley-Blackwell; 2000.
Shubin, Neil. A História de quando éramos peixes: uma revolucionária teoria sobre a origem do corpo humano editora. 1ª edição Rio de Janeiro: Elsevier, 2008. 191 p.
Freitas, R. et al. Hoxd13 Contribution to the Evolution of Vertebrate Appendages Developmental Cell - 11 December 2012 (Vol. 23, Issue 6, pp. 1219-1229. doi: 10.1016/j.devcel.2012.10.015
Schneider, Igor, Shubin, Neil H. Making Limbs from Fins Developmental Cell - 11 December 2012 Vol. 23, Issue 6, pp. 1121-1122. doi: 10.1016/j.devcel.2012.11.011
Vogel G. Developmental biology. Turing pattern fingered for digit formation. Science. 2012 Dec 14;338(6113):1406. doi: 10.1126/science.338.6113.1406. PubMed PMID: 23239707.
Sheth R, Marcon L, Bastida MF, Junco M, Quintana L, Dahn R, Kmita M, Sharpe J, Ros MA. Hox genes regulate digit patterning by controlling the wavelength of a Turing-type mechanism. Science. 2012 Dec 14;338(6113):1476-80. doi:10.1126/science.1226804. PubMed PMID: 23239739.
Boraas, ME, Seale, DB, & Boxhorn, JE (1998). Phagotrophy by a flagellate selects for colonial prey: A possible origin of multicellularity. Evolutionary Ecology Volume 12, Issue 2 , pp 153-164. DOI: 10.1023/A:1006527528063
Miller, S. M. (2010) Volvox, Chlamydomonas, and the Evolution of Multicellularity. Nature Education 3(9):65 [Link]
Ratcliff, W.C., J. Pentz and M. Travisano. 2013. Tempo and mode of multicellular adaptation in experimentally-evolved Saccharomyces cerevisiae. Evolution. 67: 1573-1581.
Ratcliff, W.C., R. F. Denison, M. Borrello and M. Travisano. 2012. Experimental evolution of multicellularity. PNAS 109:5, 1595-1600.
Miller, A. J. 2007. Crop Plants: Evolution. eLS.
Drake, Abby Grace and Klingenberg, Christian Peter Large‐Scale Diversification of Skull Shape in Domestic Dogs: Disparity and Modularity Am Nat 2010. Vol. 175, pp. 289-301 DOI: 10.1086/650372
Wayne, Robert K. (1986). “Cranial Morphology of Domestic and Wild Canids: The Influence of Development on Morphological Change.” Evolution, Vol. 40, No. 2 (Mar., 1986), pp. 243-261 * Stable URL: http://www.jstor.org/stable/2408805
Felsenstein, J. 1985. Phylogenies and the comparative method. American Naturalist 125:1-15.
Diniz-Filho JAF (2000). Métodos Filogenéticos Comparativos Holos Editora. Ribeirão Preto
Hernández, C. E., Rodríguez-Serrano, E., Avaria-Llautureo, J., Inostroza-Michael, O., Morales-Pallero, B., Boric-Bargetto, D., Canales-Aguirre, C. B., Marquet, P. A., Meade, A. 2013, Using phylogenetic information and the comparative method to evaluate hypotheses in macroecology. Methods in Ecology and Evolution, 4: 401–415. doi: 10.1111/2041-210X.12033
Lenski, R.E., Ofria,C., Robert T. Pennock, Christoph Adami The Evolutionary Origin of Complex Features Nature Vol. 423, 2003, pp. 139-145.
O'Neill B (2003) Digital Evolution. PLoS Biol 1(1): e18. doi:10.1371/journal.pbio.0000018
Wilke CO, Adami C (2002) The biology of digital organisms. Trends Ecol Evol 17: 528–532
Yaeger, L. S. 1994. Computational Genetics, Physiology, Metabolism, Neural Systems, Learning, Vision, and Behavior or PolyWorld: Life in a New Context. Langton, C. ed. Proceedings of the Artificial Life III Conference. 263-298. Addison-Wesley.
González-Candelas, Fernando, Bracho, María Alma, Wróbel, Borys and Moya, Andrés Molecular evolution in court: analysis of a large hepatitis C virus outbreak from an evolving source BMC Biology 2013, 11:76 doi:10.1186/1741-7007-11-76 - http://www.biomedcentral.com/1741-7007/11/76
Scaduto DI, Brown JM, Haaland WC, Zwickl DJ, Hillis DM, Metzker ML. Source identification in two criminal cases using phylogenetic analysis of HIV-1 DNA sequences. Proc Natl Acad Sci U S A. 2010 Dec 14;107(50):21242-7. doi: 10.1073/pnas.1015673107.
Futuyma, Douglas J. The Uses of Evolutionary Biology. Science, New Series, Vol. 267, No. 5194 (Jan. 6, 1995), 41-42.
Hendry, A. P., Kinnison, M. T., Heino, M., Day, T., Smith, T. B., Fitt, G., Bergstrom, C. T., Oakeshott, J., Jørgensen, P. S., Zalucki, M. P., Gilchrist, G., Southerton, S., Sih, A., Strauss, S., Denison, R. F. and Carroll, S. P. (2011), Evolutionary principles and their practical application. Evolutionary Applications, 4: 159–183. doi: 10.1111/j.1752-4571.2010.00165.x
Bull, J. J. and Wichman, H. A. Applied evolution Annual Review of Ecology and Systematics Vol. 32: 183-217 Volume publication date November 2001 DOI: 10.1146/annurev.ecolsys.32.081501.114020
Nesse RM. How is Darwinian medicine useful? West J Med. 2001 May;174(5):358-60. Review. PubMed PMID: 11342524; PubMed Central PMCID: PMC1071402.
Nesse RM, Ganten D, Gregory TR, Omenn GS. Evolutionary molecular medicine. J Mol Med (Berl). 2012 May;90(5):509-22. Epub 2012 Apr 29. PubMed PMID: 22544168
Nesse RM. What evolutionary biology offers public health. Bull World Health Organ. 2008 Feb;86(2):83. PubMed PMID: 18297156; PubMed Central PMCID:PMC2647384.
Nesse RM, Williams GC. Evolution and the origins of disease. Sci Am. 1998 Nov;279(5):86-93. Erratum in: Sci Am 1999 Mar;280(3):14. PubMed PMID: 9796548.
Reiber C. Evolution for epidemiologists. Ann Epidemiol. 2009 Apr;19(4):276-9. doi: 10.1016/j.annepidem.2008.12.016. Epub 2009 Feb 20. Review. PubMed PMID:19230709.
Daoust, Simon P.,Fahrig, Lenore, Martin, Amanda E.,Thomas, Frédéric . From forest and agro-ecosystems to the microecosystems of the human body: what can landscape ecology tell us about tumor growth, metastasis, and treatment options? Evolutionary Applications, 2013; 6 (1): 82 DOI: 10.1111/eva.12031
Greaves, Mel . Cancer stem cells as ‘units of selection’. Evolutionary Applications, 2013; 6 (1): 102 DOI: 10.1111/eva.12017
Nunney, Leonard The real war on cancer: the evolutionary dynamics of cancer suppression. Evolutionary Applications, 2013; 6 (1): 11 DOI: 10.1111/eva.12018
Foo, Jasmine, Leder, Kevin, Mumenthaler, Shannon M. Cancer as a moving target: understanding the composition and rebound growth kinetics of recurrent tumors. Evolutionary Applications, 2013; 6 (1): 54 DOI: 10.1111/eva.12019
---------------------------------------
Crédito das figuras:
Biólogo em campo: PASCAL GOETGHELUCK/SCIENCE PHOTO LIBRARY
Trabalho de campo geológicot: DAVID NUNUK/SCIENCE PHOTO LIBRARY
Análise de DNA fóssil: VINCENT MONCORGE/LOOK AT SCIENCES/SCIENCE PHOTO LIBRARY